Niacina
Contenido
Resumen
- Los precursores dietarios del nicotinamida adenina dinucleótido (NAD), incluyendo el ácido nicotínico, la nicotinamida, y la nicotinamida ribósido, son colectivamente referidos como niacina o vitamina B3. El aminoácido esencial triptófano puede también ser convertido en NAD a través de la vía de quinurenina. (Más información)
- El NAD puede ser fosforilado (NADP) y reducido (NADH y NADPH). El NAD funciona en las reacciones de oxidación-reducción (redox) y en las reacciones no-redox. (Más información)
- La pelagra es una enfermedad de deficiencia severa de niacina. Se caracteriza por síntomas que afectan la piel, el sistema digestivo, y el sistema nervioso; la pelagra puede causar la muerte si no es tratada. (Más información)
- Las causas de deficiencia de niacina incluyen una ingesta oral inadecuada, la escasa biodisponibilidad de granos sin nixtamalizar, la absorción defectuosa de triptófano, desordenes metabólicos, y el uso a largo plazo de tratamientos quimioterapéuticos. (Más información)
- Los requerimientos de la ingesta dietaria para la niacina están basados en la excreción urinaria de los metabolitos de niacina. (Más información)
- El NAD es el único sustrato para las enzimas PARP y sirtuinas implicadas en las actividades de reparación del ADN; por lo que NAD es crítico para la estabilidad del genoma. Varios estudios, la mayoría usando modelos in vitro y animales, sugieren un posible papel para la niacina en la prevención del cáncer. En un reciente ensayo de fase III, se encontró que una dosis farmacológica de nicotinamida redujo la tasa de lesiones premalignas de la piel y cánceres no melanoma en sujetos en alto riesgo. (Más información)
- A pesar de los resultados prometedores iniciales, la administración de nicotinamida ha fallado en prevenir o retrasar la aparición de la diabetes mellitus tipo 1 en familiares en alto riesgo de pacientes diabéticos tipo 1. Investigación futura podría explorar el uso de nicotinamida en la terapia combinada y evaluar los activadores de las enzimas dependiente de NAD. (Más información)
- En dosis farmacológicas, el ácido nicotínico mejoró los perfiles lipídicos de pacientes con un historial de enfermedad vascular, sin embargo, falló en reducir eventos cardiovasculares recurrentes o la mortalidad. (Más información)
- El desglose elevado de triptófano en la vía de quinurenina y la deficiencia de niacina han sido reportados en personas VIH positivas. Sin embargo, en la actualidad, es necesario un mejor entendimiento del papel de la vía de quinurenina y otras rutas biosintéticas del NAD durante la infección del VIH para establecer si esta población podría beneficiarse de la suplementación con niacina. (Más información)
- La mayoría de los efectos adversos de la niacina (ácido nicotínico y nicotinamida) han sido reportados con dosis farmacológicas de ácido nicotínico. El nivel máximo de ingesta tolerable (NM) para la niacina está basado en la prevención del enrojecimiento de la piel, el efecto secundario más importante de la niacina. La coadministración con laropiprant — un antagonista del receptor 1 de prostaglandina D2 — ayuda a reducir el enrojecimiento de la piel inducido por el ácido nicotínico. (Más información)
Niacina o vitamina B3 es una vitamina hidrosoluble usada por el cuerpo para formar la coenzima nicotinamida, NAD+. El termino 'niacina' es frecuentemente usado para referirse al acido nicotínico (ácido piridina-3-carboxílico) únicamente, aunque otros vitámeros con un anillo de piridina, incluyendo la nicotinamida (piridina-3-carboxamida) y nicotinamida ribósido, también contribuyen a la formación de NAD+ (1). Ninguno de los vitámeros está relacionado a la nicotina encontrada en el tabaco, aunque sus nombres sean similares. De igual manera la nicotina — pero no el ácido nicotínico — es un agonista de los receptores nicotínicos que responden al neurotransmisor, acetilcolina.
Metabolismo
Esencial para todas las formas de vida, la coenzima nicotinamida NAD+ es sintetizada en el cuerpo a partir de cuatro precursores que son proporcionados en la dieta: el acido nicotínico, la nicotinamida, nicotinamida ribósido, y el triptófano (Figura 1).
La Figura 2 ilustra las distintas rutas biosintéticas que conducen a la producción de NAD+ proveniente de varios precursores dietarios. NAD+ es sintetizada a partir de la nicotinamida y nicotinamida ribósido a través de dos reacciones enzimáticas, mientras que la ruta que produce NAD+ del ácido nicotínico — conocida como la ruta de Preiss-Handler — incluye tres pasos. La vía de quinurenina es la ruta biosintetica mas larga del NAD+: el catabolismo del triptófano a través de la quinurenina produce ácido quinolínico, el cual es entonces convertido a ácido nicotínico mononucleótido, un intermedio en el metabolismo del NAD+. NAD+ es entonces sintetizado a partir del ácido nicotínico mononucleótido en la ruta de Preiss-Handler (2).
Todas las rutas generan mononucleótidos intermediarios — ácido nicotínico mononucleótido o nicotinamida mononucleótido. Enzimas específicas, conocidas como fosforribosiltransferasas, catalizan la adición de un grupo fosforribosa al ácido nicotínico o ácido quinolínico para producir ácido nicotínico mononucleótido o en la nicotinamida para generar nicotinamida mononucleótido. El nicotinamida mononucleótido también es generado por la fosforilación de nicotinamida ribósido, catalizada por quinasas nicotinamida ribosa (NRK). Además, las adenililtransferasas catalizan la adenilación de estos mononucleótidos para formar ya sea ácido nicotínico adenina dinucleótido o NAD+. El ácido nicotínico adenina dinucleótido es luego convertido a NAD+ por la NAD+ sintetasa dependiente de glutamina (NADSYN), la cual usa glutamina con un donante de grupo amida (Figura 2) (2). Debe notarse que se ha reportado que el ácido nicotínico adenina dinucleótido se forma después de la administración de nicotinamida ribósido en alta dosis, sugiriendo que una potencial desamidación podría ocurrir para convertir el NAD+ a ácido nicotínico adenina dinucleótido cuando el depósito de NAD+ es alto (1).
La NAD quinasa cataliza la fosforilación del NAD a NADP usando adenosil trifosfato (ATP) como donador de fosforilo (3). Las propiedades de oxido-reducción (redox) del dinucleótido no son afectadas por la fosforilación tal que las parejas redox NAD+/NADH y NADP+/NADPH muestran potenciales redox similares (4). La oxidación y reducción de la posición C-4 del grupo nicotinamida del NAD y su forma fosforilada son esenciales para las reacciones de transferencia de electrones que soportan funciones metabólicas y bioenergéticas vitales en todas las células (véase Función). Así, NAD y NADP son reciclados una y otra vez entre la forma oxidada (NAD+ y NADP+) y reducida, como se muestra en Figura 3.

[Figura 1 - Clic para Agrandar]

[Figura 2 - Clic para Agrandar]
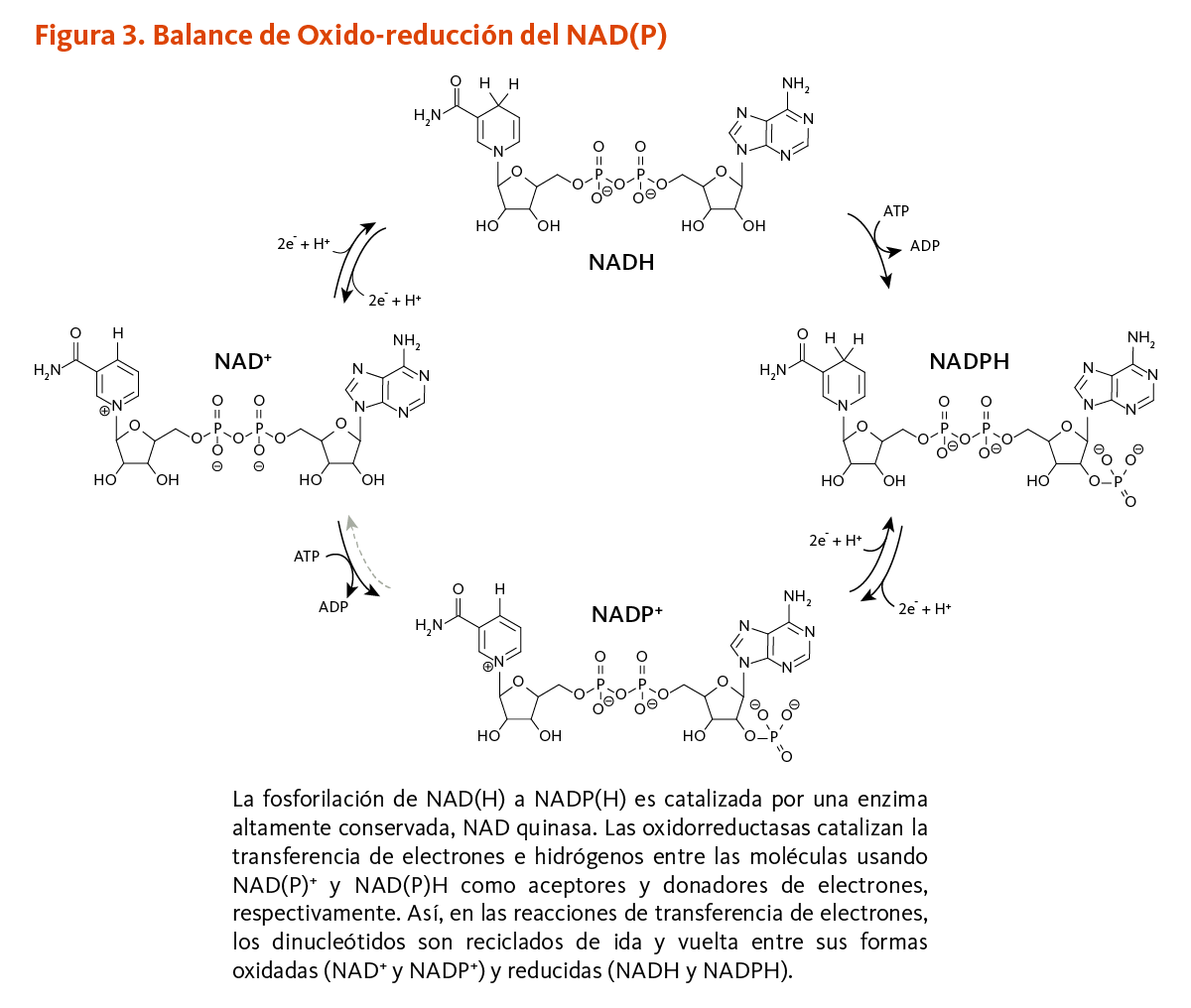
[Figura 3 - Clic para Agrandar]
Función
NAD como una coenzima en las reacciones de transferencia de electrones
Los organismos vivos derivan la mayoría de su energía de reacciones redox, procesos que involucran la transferencia de electrones. Mas de 400 enzimas requieren las coenzimas de niacina, NAD y NADP, principalmente para aceptar o donar electrones para las reacciones redox (5). NAD y NADP parecen soportar funciones distintas (Figura 4). NAD funciona más frecuentemente en reacciones productoras de energía que involucran la degradación (catabolismo) de carbohidratos, grasas, proteínas, y alcohol. NADP generalmente sirve en las reacciones biosintéticas (anabolismo), como en la síntesis de ácidos grasos, esteroides (por ejemplo, colesterol, ácidos biliares y hormonas esteroides), y como bloques de construcción de otras macromoléculas (4). NADP es también esencial para la regeneración de componentes de la detoxificación y de sistemas antioxidantes (4). Para soportar estas funciones, la célula mantiene NAD en un estado ampliamente oxidado (NAD+) para que sirva como un agente oxidante en las reacciones catabólicas, mientras que NADP se mantiene en gran medida en un estado reducido (NADPH) para donar fácilmente electrones para los procesos celulares reductivos (4, 6).
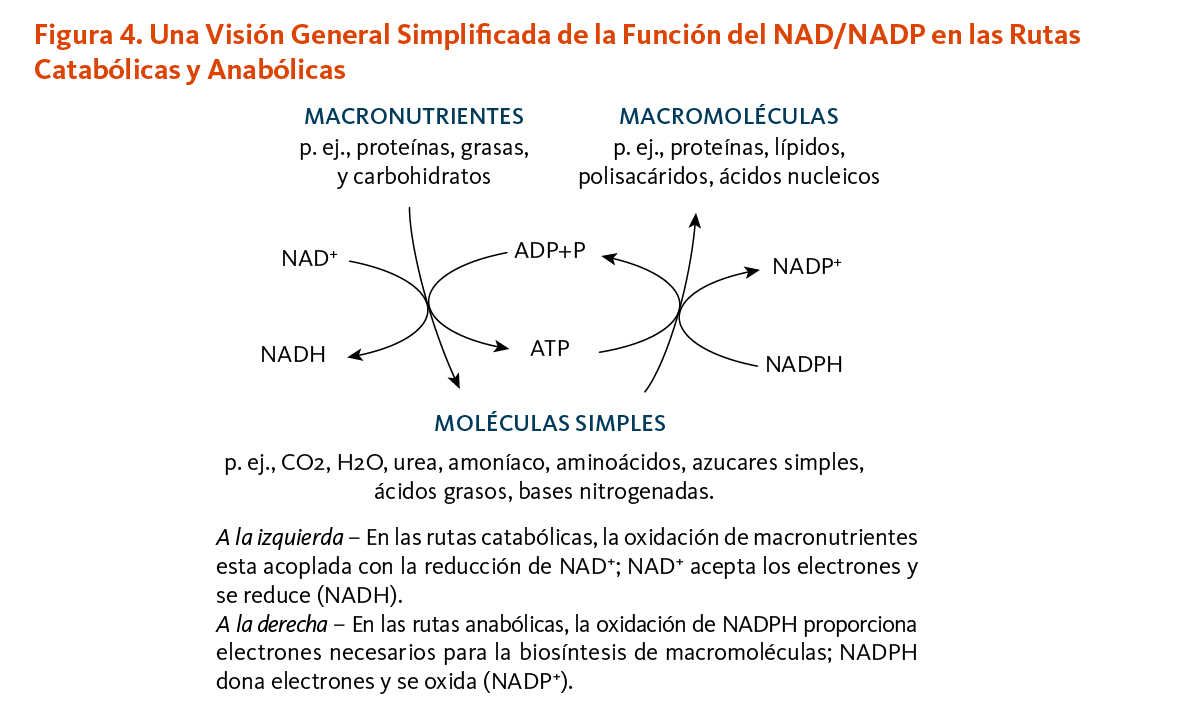
[Figura 4 - Clic para Agrandar]
NAD como un sustrato para las enzimas que consumen NAD
La coenzima de la niacina, NAD, es el sustrato (reactante) de por lo menos cuatros clases de enzimas. Dos clases de enzimas con actividades de mono adenosina difosfato (ADP)- ribosiltransferasa y/o poli (ADP-ribosa) polimerasa catalizan las reacciones de transferencia de ADP-ribosilo. Proteínas parecidas al regulador de información silencioso 2 (Sir2) (sirtuinas) catalizan la eliminación de los grupos acetilo provenientes de proteínas acetiladas, utilizando ADP-ribosa del NAD como un como un aceptor para grupos de acetilo. Finalmente, las ADP-ribosil ciclasas están involucradas en la regulación de la señalización del calcio intracelular.
ADP-ribosilación
Las enzimas con actividades ADP-ribosiltransferasa estaban previamente divididas entre mono ADP-ribosiltransferasas (ART) y poli (ADP-ribosa) polimerasas (PARP). Las ART fueron primeramente descubiertas en ciertas bacterias patogénicas — como aquellas causantes del cólera y difteria — donde median las acciones de toxinas. Estas enzimas transfieren un grupo de residuo ADP-ribosa del NAD a un amino acido especifico de una proteína objetivo, con la creación de una proteína ADP-ribosilada y la liberación de nicotinamida.
Debido a que se ha encontrado que la mayoría de las PARP exhiben solo actividades mono ADP-ribosiltransferasa una nueva nomenclatura fue propuesta para las enzimas que catalizan la ADP-ribosilación: Una familia de mono ADP-ribosiltransferasas con homología con toxinas diftéricas bacterianas fue llamada ARTD, mientras que las enzimas con actividades mono o poli ADP-ribosiltransferasa y relacionadas a las toxinas clostridiales C2 y C3 fueron incluidas en la familia ARTC (7, 8).
ARTC son enzimas extracelulares que catalizan la mono ADP-ribosilación de proteínas secretadas o de membrana involucradas en la inmunidad innata y comunicación celular (2).
ARTD son enzimas intracelulares con actividades mono o poli ADP-ribosiltransferasa. Por lo menos 18 ARTD han sido identificadas. Todas las ARTD poseen un dominio catalítico similar a la toxina diftérica que se une a NAD+. Solo las ARTD 1, 2, 5 y 6 catalizan transferencias poli (ADP-ribosa); las otras tienen actividades mono ADP-ribosiltransferasas. Se demostró que las ARTD están involucradas en la reparación del ADN y respuestas al estrés, señalización celular, regulación de la transcripción, apoptosis, diferenciación celular, mantenimiento de la integridad genómica, y defensa antiviral (revisado en 8).
Desacetilación dependiente de NAD
Siete sirtuinas (SIRT 1-7) han sido identificadas en humanos. Las sirtuinas son una clase de enzimas deacetilasa dependientes de NAD que remueven grupos de acetilo de los restos de lisina acetilados de proteínas diana. Durante el proceso de desacetilación, el grupo acetilo es transferido al grupo ADP-ribosa escindido del NAD produciendo O-acetil-ADP-ribosa. La nicotinamida puede ejercer una inhibición de retroalimentación en la reacción de desacetilación (9). Al igual que la ADP-ribosilación, la acetilación es una modificación postraduccional que afecta la función de las proteínas diana. El interés inicial en las sirtuinas ocurrió después del descubrimiento de que su activación podía imitar la restricción calórica, la cual se ha mostrado incrementa la esperanza de vida en organismos inferiores. Tal papel en los mamíferos es controversial, aunque las sirtuinas son reguladores de detección de energía involucrados en las rutas de señalización que podrían jugar papeles importantes en el retraso de la aparición de enfermedades relacionadas a la edad (p. ej., enfermedades cardiovasculares, cáncer, demencia, artritis). Hasta la fecha, el espectro de sus funciones biológicas incluye el silenciamiento de genes, la reparación del daño al ADN, la regulación del ciclo celular, y la diferenciación celular.
Movilización del calcio
En los humanos CD38 y CD157 pertenecen a una familia de NAD+glicohidrolasas/ADP-ribosil ciclasas. Estas enzimas catalizan la formación de reguladores clave de la señalización por calcio, es decir ADP-ribosa (lineal), ADP-ribosa cíclica, y ácido nicotínico adenina dinucleótido fosfato (Figura 5). El ADP-ribosa cíclica y el ácido nicotínico adenina dinucleótido fosfato trabajan dentro de las células para provocar la liberación de los iones de calcio de los sitios de almacenamiento internos (es decir, el retículo endoplásmico, lisosomas, mitocondrias), mientras que el ADP-ribosa estimula la entrada de calcio extracelular a través de los canales de cationes TRPM2 de la membrana celular (2). Otro agonista de, TRPM2, 2’-desoxi-ADP-ribosa, fue identificado recientemente in vitro. Se encontró que CD38 cataliza la síntesis de 2’-desoxi-ADP-ribosa a partir del nicotinamida mononucleótido y 2’-desoxi-ATP (11). O-acetil-ADP-ribosa generada por la actividad de las sirtuinas también controla la entrada de calcio a través de los canales TRPM2 (6). La transducción de señal mediada por calcio intracelular es regulada por la entrada transitoria de calcio en la célula o la liberación de calcio de almacenamientos intracelulares. La señalización por calcio esta críticamente involucrada en procesos como la neurotransmisión, la liberación de insulina de células β pancreáticas, la contracción muscular, y la activación de linfocitos-T (6).
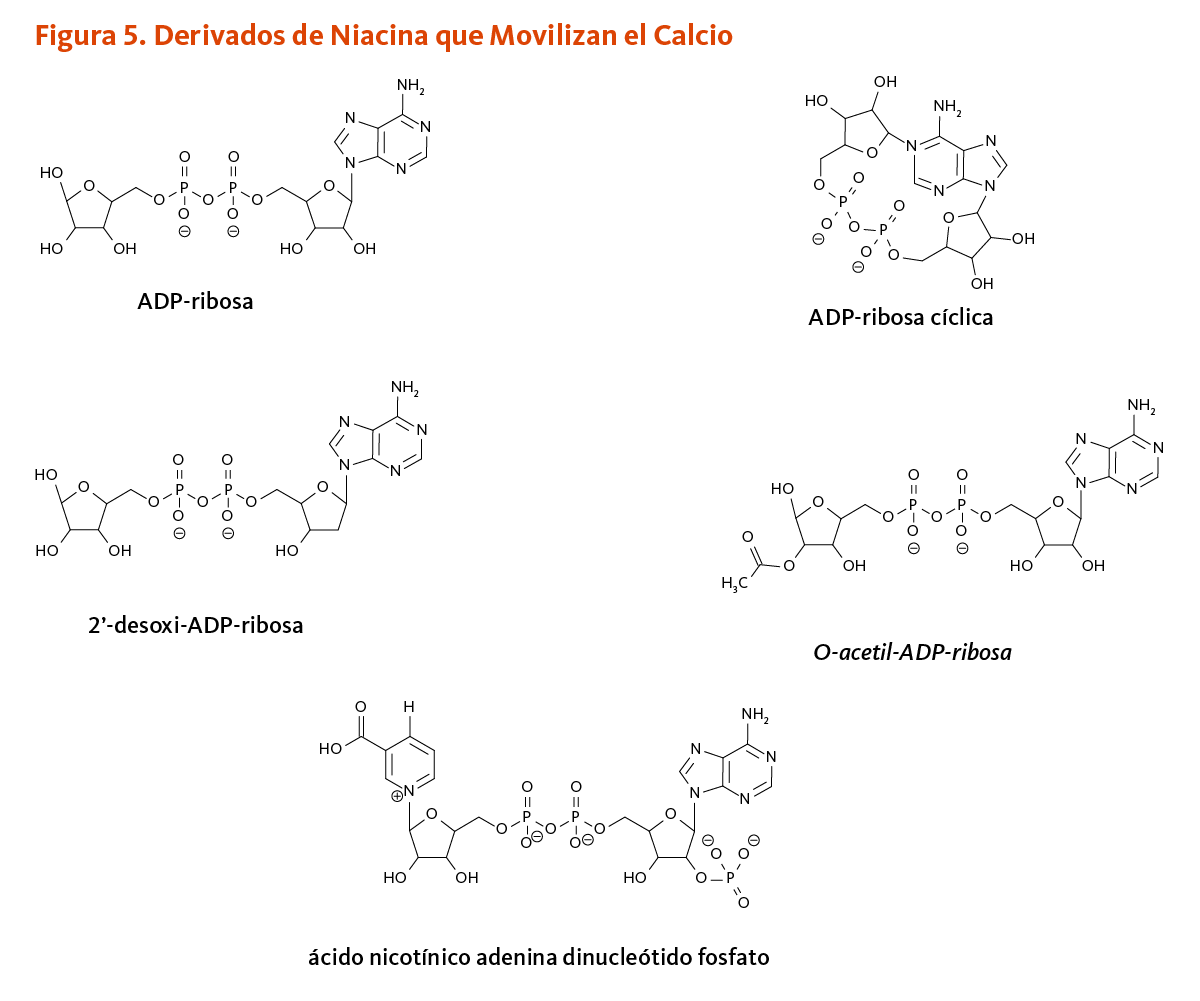
[Figura 5 - Clic para Agrandar]
NAD como un ligando
NAD+ ha sido identificado como un agonista endógeno de receptores de membrana purinérgicos de subclase P2Y. en particular, se encontró que NAD se une al receptor P2Y1 y actúa como un neurotransmisor inhibitorio en las uniones neuromusculares en los músculos lisos viscerales (12). También se encontró que el NAD+ extracelular se comporta como una citoquina proinflamatoria, desencadenando la activación de granulocitos aislados. La unión de NAD+ al receptor P2Y11 en la superficie del granulocito activa una cascada de señalización involucrando la ADP-ribosa cíclica y el aumento del calcio intracelular, eventualmente estimulando la generación de superóxido y la quimiotaxis (13). Observaciones similares fueron hechas con monolitos activados por lipopolisacáridos (14). NAADP+ extracelular y ADP-ribosa podrían también unirse a los receptores P2Y y desencadenar NAADP+ intracelular — y ADP-ribosa-dependiente de la movilización del calcio (véase Movilización del calcio) (15, 16).
Efectos hipolipemiantes con dosis farmacológicas de ácido nicotínico
Por más de medio siglo, las dosis farmacológicas de ácido nicotínico, pero no de nicotinamida, han sido conocidas por reducir el colesterol del suero (véase Tratamiento de Enfermedades) (17). Sin embargo, los mecanismos exactos que subyacen el efecto hipolipemiante del ácido nicotínico siguen siendo especulativos. Dos receptores de membrana acoplados a proteína G, GPR109A y GPR109B, se unen al acido nicotínico con alta y baja afinidad, respectivamente. Estos receptores de ácido nicotínico son principalmente expresados en el tejido adiposo y células inmunes (pero no los linfocitos). También son encontrados en las células epiteliales colónicas y pigmentadas de la retina, queratinocitos, células mamarias, microglía, y posiblemente en niveles bajos en el hígado (18). Así, los efectos modificadores de lípidos del ácido nicotínico son mas propensos a ser mediados por mecanismos independientes de receptores en tejidos principales del metabolismo de los lípidos como el hígado y el musculo esquelético. Datos in vitro tempranos sugirieron que el ácido nicotínico podría perjudicar la secreción de lipoproteínas de muy baja densidad (VLDL) al inhibir la síntesis de triglicéridos y desencadenar la degradación de lipoproteína ApoB en los hepatocitos (19). En otro estudio, el ácido nicotínico afecto la absorción hepática de lipoproteínas ApoAI, reduciendo así la eliminación de lipoproteínas de alta densidad (HDL) de la circulación (revisado en 20). En los adipocitos se encontró que la unión del acido nicotínico al GPR109A inicio una cascada de transducción de señales resultando en reducciones en la producción de ácidos grasos libres mediante la inhibición de la lipasa sensible a las hormonas involucrada en la lipólisis de triglicéridos (21). Sin embargo, observaciones recientes han sugerido que el efecto hipolipemiante del ácido nicotínico no se debe a su actividad anti-lipolítica (22). Ensayos mostraron que los agonistas sintéticos del GPR109A agudamente disminuyeron los ácidos grasos libres, pero fallaron en afectar los lípidos del suero (22). Aparte de su impacto en los HDL y otros lípidos del plasma, el ácido nicotínico ha exhibido actividades anti-ateroscleróticas en monocitos, macrófagos, o células endoteliales vasculares cultivadas, al modular la inflamación y estrés oxidativo y regulando la adhesión, migración, y diferenciación celular (revisado en 18).
Deficiencia
Pelagra
Causas
La última etapa de la deficiencia severa de niacina se conoce como pelagra. Los primeros registros de pelagra ocurrieron luego del cultivo generalizado de maíz en Europa en 1700 (23). La enfermedad es generalmente asociada con las clases sociales más pobres en la cuales el alimento básico principal consistía en cereales como el maíz o el sorgo. La pelagra también fue común en el sur de los Estados Unidos durante los inicios del año 1900, donde el ingreso era bajo y los productos de maíz eran un alimento básico importante (24). Curiosamente, la pelagra no era conocida en México, donde el maíz era también un alimento básico importante y mucha de la población era también pobre. De hecho, si el maíz contiene cantidades considerables de niacina, esta se encuentra presente en una forma ligada que no está nutricionalmente disponible para los seres humanos. La preparación tradicional de tortillas de maíz en México involucra remojar el maíz en una solución de cal (óxido de calcio) antes de cocinarlo. Calentar el maíz en una solución alcalina produce la liberación de la niacina ligada, aumentando así su biodisponibilidad (25). La epidemia de pelagra fue también desconocida por los nativos americanos quienes consumían maíz inmaduro que contiene predominantemente niacina desligada (biodisponible) (24).
La deficiencia de niacina o pelagra pueden resultar de la ingesta dietaría inadecuada de precursores de NAD, incluyendo triptófano. La deficiencia de niacina — frecuentemente asociada con la malnutrición — se observa en la población sin hogar, en individuos que sufren de anorexia nerviosa u obesidad, y en consumidores de dietas altas en maíz y pobres en proteína animal (26-29). Deficiencias de otras vitaminas B y algunos minerales traza pueden agravar la deficiencia de niacina (30, 31). Trastornos de malabsorción que pueden conducir a la pelagra incluyen la enfermedad de Crohn y megaduodeno (32, 33). Pacientes con la enfermedad de Hartnup, un trastorno hereditario que resulta en una absorción defectuosa de triptófano, han desarrollado pelagra (véase Trastornos genéticos sensibles a la niacina). El síndrome carcinoide, una condición de aumento de la secreción de serotonina y otras catecolaminas por tumores carcinoides, puede también resultar en pelagra debido al incremento en la utilización de triptófano para la serotonina en lugar de la síntesis de niacina. Además, el tratamiento prolongado con el fármaco anti-tuberculosis isoniacida ha resultado en deficiencia de niacina (34). Otros agentes farmacéuticos, incluyendo las drogas inmunosupresoras azatioprina (Imuran) y 6- mercaptopurina, el fármaco contra el cáncer 5-fluorouracilo (5-FU, Adrucil), y levodopa/carbidopa (Sinemet; dos fármacos dados a personas con la enfermedad de Parkinson) son conocidos por incrementar la dependencia en la niacina dietaría al interferir con la vía triptófano-quinurenina-niacina (35). Finalmente, otras poblaciones en riesgo de una deficiencia de niacina incluyen pacientes en diálisis, pacientes con cáncer (36, 37), individuos que sufren de alcoholismo crónico (38), y personas con VIH (véase VIH/SIDA abajo). Además, la ingesta crónica de alcohol puede llevar a una deficiencia de niacina severa a través de la reducción de la ingesta de niacina dietaría y al interferir con la conversión de triptófano-a-NAD (30).
Síntomas de la deficiencia
Los síntomas más comunes de la deficiencia de niacina involucran a la piel, al sistema digestivo, y al sistema nervioso. Los síntomas de pelagra son comúnmente referidos como a las tres D: dermatitis sensible al sol, diarrea, y demencia. Una cuarta D, deceso, ocurre si la pelagra no es tratada (5). En la piel, un sarpullido de pigmentación oscura, grueso y escamoso, se desarrolla simétricamente en las áreas expuestas a la luz del sol. De hecho, la palabra "pelagra" deriva de "pelle agra" la expresión italiana para la piel áspera. Los síntomas relacionados al sistema digestivo incluyen inflamación de la boca y lengua ("lengua roja brillante"), vómitos, constipación, dolor abdominal y por último diarrea. Los trastornos gastrointestinales y la diarrea contribuyen a la desnutrición continua de los pacientes. Los síntomas neurológicos incluyen dolor de cabeza, apatía, fatiga, depresión, desorientación, y pérdida de memoria y son más consistentes con delirio que con la demencia descrita históricamente (38). La presentación de la enfermedad varía en apariencia ya que la triada clásica raramente se presenta en su totalidad. La ausencia de la dermatitis, por ejemplo, es conocida como pelagra sinusoidal pelagra.
Tratamiento
Para tratar la pelagra, la Organización Mundial de la Salud (OMS) recomienda administrar nicotinamida para prevenir el enrojecimiento comúnmente causado por el ácido nicotínico (véase Seguridad). Los lineamientos del tratamiento sugieren usar 300 mg/día de nicotinamida oral en dosis divididas, o 100 mg/día administrados parenteralmente en dosis divididas, por tres a cuatro semanas (37, 39). Debido a que los pacientes con pelagra frecuentemente muestran deficiencias vitamínicas adicionales, la administración de una preparación de complejo de vitamina B es aconsejada (39).
La Ingesta Diaria Recomendada (IDR)
Equivalente de niacina (EN)
El termino "equivalente de niacina" (EN) es usado para describir la contribución de la ingesta dietaría de todas las formas de niacina que están disponibles para el cuerpo. En los individuos sanos, menos del 2% del triptófano dietario es convertido a NAD en la vía de quinurenina (40). La síntesis del NAD a partir del triptófano es bastante ineficiente y depende de enzimas que requieren de vitamina B6 y riboflavina como también de un enzima que contiene hemo (hierro). Sin embargo, el triptófano es esencial como precursor para el NAD+. Defectos congénitos en el trasporte del triptófano y el metabolismo resultan en trastornos clínicos severos atribuidos al agotamiento de NAD+ (véase Trastornos genéticos sensibles a la niacina). En promedio, 60 miligramos (mg) de triptófano se considera corresponden a 1 mg de niacina o 1 mg de EN.
Ingestas diarias recomendadas
La ingesta diaria recomendada (IDR) para la niacina esta basada en la prevención de la deficiencia. La pelagra puede ser prevenida con alrededor de 11 mg de EN/día, pero se ha encontrado que, de 12 mg a 16 mg de EN/día, se normaliza la excreción urinaria de metabolitos de niacina (productos de degradación) en adultos jóvenes sanos. Debido a que la pelagra representa una deficiencia severa, la Junta de Nutrición y Alimentos (JNA) del Instituto de Medicina de EE.UU. optó por utilizar la excreción de metabolitos de niacina como un indicador del estado nutricional de la niacina más que como un indicador de síntomas de pelagra (41). Sin embargo, se ha argumentado que el contenido de NAD y NADP celulares pueden ser indicadores más relevantes del estatus nutricional de la niacina (24).
| Etapa de la Vida | Edad | Machos (mg EN*/día) | Hembras (mg EN/día) |
|---|---|---|---|
| Infantes | 0-6 meses | 2 (IA) | 2 (IA) |
| Infantes | 7-12 meses | 4 (IA) | 4 (IA) |
| Niños | 1-3 años | 6 | 6 |
| Niños | 4-8 años | 8 | 8 |
| Niños | 9-13 años | 12 | 12 |
| Adolescentes | 14-18 años | 16 | 14 |
| Adultos | 19 años y más | 16 | 14 |
| Embarazo | Todas las edades | - | 18 |
| Período de lactancia | Todas las edades | - | 17 |
| *EN, equivalentes de niacina: 1 mg de EN = 60 mg de triptófano = 1 mg de niacina | |||
Prevención de Enfermedades
Cáncer
Estudios de cultivos celulares (in vitro) aportan evidencia de que el contenido de NAD influencia mecanismos que mantienen la estabilidad genómica. La pérdida de estabilidad genómica, caracterizada por una alta tasa de daño en el ADN y cromosomas, es un sello distintivo del cáncer (42). La comprensión actual es que se reduce la piscina de NAD durante la deficiencia de niacina y esto afecta la actividad de enzimas que consumen NAD en lugar de funciones redox y metabólicas (43). Entre las reacciones dependientes de NAD, poli ADP-ribosilaciones catalizadas por enzimas PARP (ARTD) son críticas para la respuesta celular al daño del ADN. Después del daño al ADN, las PARP son activadas; se mostró que las subsiguientes poli ADP-ribosilaciones de una serie de señalización y moléculas estructurales por las PARP facilitan la reparación del ADN en roturas de la de cadena del ADN (44). Se ha encontrado que el agotamiento celular de NAD disminuye los niveles de la proteína supresora de tumores, p53, un objetivo para la poli ADP-ribosilación en células de mama, piel y pulmón humanas (45). La expresión de p53 fue también alterada por la deficiencia de niacina en las células de la medula ósea de ratas (46). El deterioro de la reparación del ADN causado por la deficiencia de niacina pudo conducir a la inestabilidad genómica e impulsar el desarrollo de tumores en modelos de ratas (47, 48). Ambas PARP y las sirtuinas han participado recientemente en el mantenimiento de la heterocromatina, un domino cromosómico asociado con la estabilidad del genoma, así como en el silenciamiento génico transcripcional, la integridad de los telómeros, y la segregación cromosómica durante la división celular (49, 50). Ni el contenido celular de NAD ni la ingesta dietaría de precursores de NAD necesarios para optimizar las respuestas de protección luego de un daño al ADN han sido determinados, pero ambos son posiblemente más altos que lo requerido para la prevención de la pelagra.
Medula ósea
Los pacientes con cáncer frecuentemente sufren de supresión de la medula ósea luego de la quimioterapia, dado que la medula ósea es uno de los tejidos mas prolíferos en el cuerpo y por lo tanto un objetivo principal para los agentes quimioterapéuticos. Se encontró que una deficiencia de niacina disminuye los niveles de NAD y poli-ADP-ribosa en la médula ósea y aumenta el riesgo de leucemia en ratas (51). Por el contrario, una dosis farmacológica de ácido nicotínico o nicotinamida fue capaz de aumentar el NAD y poli-ADP-ribosa en la medula ósea y disminuir el desarrollo de la leucemia en ratas (52). Se ha sugerido que la deficiencia de niacina a menudo observada en pacientes con cáncer podría sensibilizar el tejido de la medula ósea al efecto supresor de la quimioterapia. Sin embargo, poco se sabe acerca de los niveles de NAD celular y la prevención del daño al ADN o del cáncer en humanos. Un estudio en dos individuos sanos involucró el aumento de los niveles de NAD en leucocitos sanguíneos a través de la suplementación con 100 mg/día de ácido nicotínico por ocho semanas. En comparación con individuos no suplementados, los individuos con suplementación experimentaron una disminución en la ruptura de hebras del ADN en los linfocitos expuestos a radicales libres en un ensayo de tubo de ensayo (53). Sin embargo, la suplementación con ácido nicotínico de hasta 100 mg/día por 14 semanas en 21 fumadores sanos falló en aportar alguna evidencia de una disminución en el daño genético inducido por el humo del cigarrillo en linfocitos sanguíneos, en comparación con el placebo (38). Más reciente, se utilizó la frecuencia de translocación cromosómica para evaluar daños al ADN en los linfocitos de sangre periférica de 82 pilotos crónicamente expuestos a la radiación ionizante, un conocido carcinógeno humano. En este estudio basado en la observación, la tasa de aberraciones cromosómicas fue significativamente menor en los sujetos con una ingesta más alta de niacina dietaría (28.4 mg/día) en comparación con una más baja (20.5 mg/día) (55). La disponibilidad mas alta de NAD+ en linfocitos de sangre periférica irradiados con rayos X se encontró favorece la reparación del ADN al mejorar la supervivencia, particularmente a través de la deacetilación de p53 mediada por SIRT (56).
Tracto digestivo superior
Generalmente, las relaciones entre los factores dietarios y el cáncer son establecidos primero en los estudios epidemiológicos y seguidos luego por investigación básica del cáncer a nivel celular. En el caso de la niacina, la investigación sobre aspectos bioquímicos y celulares de la reparación del ADN ha estimulado un interés en la relación entre la ingesta de niacina y el riesgo de cáncer en la población humana (57). Un estudio de caso y control de gran magnitud encontró que el consumo aumentado de niacina, junto con nutrientes antioxidantes, se asociaba con una incidencia disminuida de cáncer oral (boca), faríngeo (garganta), y esofágico en el norte de Italia y Suiza. Un aumento de la ingesta de niacina diaria de 6.2 mg se asoció con un descenso de alrededor del 40% de los casos de cáncer de boca y garganta, mientras un aumento de 5.2 mg en la ingesta diaria de niacina se asoció con un descenso similar en los casos de cáncer esofágico (58, 59).
Piel
La deficiencia de niacina puede llevar a una sensibilidad severa a la luz solar en la piel expuesta. Dada la implicación de las enzimas dependientes de NAD en la reparación del ADN, ha habido cierto interés en el efecto de la niacina en la salud de la piel. Experimentos in vitro y en animales han ayudado a obtener información, pero los datos en humanos sobre el estatus de la niacina/NAD y el cáncer de piel son muy limitados. Un estudio reportó que la suplementación con niacina disminuyó el riesgo de canceres en la piel inducidos por luz ultravioleta (UV) en ratones, a pesar del hecho de que los ratones convierten triptófano a NAD más eficientemente que las ratas y los humanos y que por lo tanto no se ven severamente deficientes (60). La hiperproliferación y la diferenciación alterada de células de la piel pueden alterar la integridad de la barrera cutánea y aumentar la ocurrencia de enfermedades de la piel pre-malignas y malignas. Un efecto protector de la niacina fue sugerido a través de la aplicación tópica de miristil nicotinato, un derivado de niacina, que aumentó con éxito la expresión de marcadores de diferenciación epidérmica en sujetos con piel fotodañada (61). La activación de receptores del ácido nicotínico, GPR109A y GPR109B, por dosis farmacológicas de niacina podría estar involucrada en la mejora de la función de la barrera cutánea. Por lo contrario, los defectos de la diferenciación en células cancerígenas cutáneas fueron ligadas a la localización celular anormal de receptores del ácido nicotínico defectuosos (62). La restricción de nicotinamida con el subsecuente agotamiento de NAD celular ha demostrado incrementar el daño del ADN inducido por el estrés oxidativo en un modelo de células precancerosas de la piel, implicando un papel protector de las vías dependientes de NAD en el cáncer (63). La disponibilidad alterada del NAD también afecta la expresión y la actividad de las sirtuinas en las células de la piel humana expuestas a UV. Junto con las PARP, las sirtuinas que consumen NAD podrían jugar un papel importante en la respuesta celular al fotodaño y la homeostasis de la piel (64).
Un análisis agrupado de dos estudios de cohorte prospectivos de gran magnitud de los EE.UU. que dio seguimiento a 41,808 hombres y 72,308 mujeres por hasta 26 años sugirió que la ingesta mas alta de niacina versus la mas baja (proveniente de la dieta y suplementos) podría proteger contra el carcinoma de células escamosas pero no contra el carcinoma de células basales y melanoma (65). Un ensayo aleatorio, doble ciego, fase III, controlado con placebo en 386 sujetos con un historial de cáncer de piel no melanoma recientemente examinó el efecto de la suplementación diaria con nicotinamida (1 g) por 12 meses en la recurrencia del cáncer de piel en intervalos de tres meses sobre un periodo de 18 meses (66). La nicotinamida efectivamente redujo la tasa de queratosis actínica premaligna (-11%), carcinoma de células escamosas (-30%) y carcinoma de células basales (-20%) en comparación al placebo después de 12 meses, sin embargo, esta protección no fue sostenida durante el periodo de seis meses posterior a la suplementación (66). Ensayos de mayor magnitud son necesarios para evaluar si la nicotinamida podría reducir el riesgo de melanomas, los cuales no son tan comunes como otros canceres de piel, pero son más mortales (67).
Diabetes mellitus tipo 1
La diabetes mellitus tipo 1 en niños es causada por la destrucción autoinmune de las células beta secretoras de insulina en el páncreas. Antes del inicio de una diabetes sintomática, anticuerpos específicos, incluyendo autoanticuerpos de las células de los islotes (ACI), pueden ser detectados en la sangre de individuos en alto riesgo (68). En un modelo experimental animal de la diabetes, niveles altos de nicotinamida son administrados para proteger las células β del daño causado por la estreptozocina (69).
Sin embargo, no se ha encontrado que dosis farmacológicas de nicotinamida (de hasta 3 g/día) sean efectivas en el retraso o prevención de la aparición de la diabetes tipo 1 en sujetos en alto riesgo. Un análisis de 10 ensayos, de los cuales cinco fueron controlados con placebo, encontró evidencia de una función de células β mejorada después de un año de tratamiento con nicotinamida, pero el análisis falló en encontrar alguna evidencia clínica de un control glucémico mejorado (70). Un ensayo controlado aleatorio multicéntrico grande de la nicotinamida en familiares ACI positivos (edades, 3-12 años) de pacientes diabéticos tipo 1 también falló en encontrar una diferencia en la incidencia de diabetes tipo 1 después de tres años (70). Un ensayo multicéntrico aleatorio, doble ciego, controlado con placebo de la nicotinamida (un máximo de 3 g/día) fue conducido en 552 familiares ACI positivos de pacientes con diabetes tipo 1. La proporción de familiares que desarrollaron diabetes tipo 1 dentro del periodo de cinco años fue comparable ya fuera si fueron tratados con nicotinamida o placebo (71). La nicotinamida pudo reducir los parámetros relacionados a la inflamación en estos sujetos en alto riesgo, pero fue ineficaz para prevenir el inicio de la enfermedad (72). Mas recientemente, reportes de caso del uso combinado de nicotinamida (25 mg/kg/día) y acetil-L-carnitina (50 mg/kg/día) en niños en riesgo de diabetes tipo 1 mostró resultados prometedores, lo que justifica una mayor investigación (73).
Tratamiento de Enfermedades
Los suplementos con niacina en dosis farmacológicas (es decir, dosis más altas que aquellas necesarias para prevenir la deficiencia) han sido usados en un intento por tratar un cierto rango de condiciones, algunas de las cuales son discutidas a continuación.
Trastornos genéticos sensibles a la niacina
Trastornos congénitos relacionados a la deficiencia de NAD pueden resultar de mutaciones en genes involucrados en la absorción y transporte de varios precursores dietarios del NAD+ o en las distintas rutas metabólicas que conllevan a la producción de NAD+ (véase Metabolismo). Algunos de estos trastornos podrían responder a la suplementación con niacina. Por ejemplo, el transporte alterado de triptófano a las células resulta en la enfermedad de Hartnup, la cual presenta signos de deficiencia grave de niacina (74). La enfermedad de Hartnup se debe a mutaciones en el gen SLCA19, el cual codifica para un transportador de aminoácidos neutros dependiente de sodio principalmente expresado en los riñones e intestinos. El manejo de la enfermedad involucra la suplementación con ácido nicotínico o nicotinamida (75). Mutaciones recesivas en genes que codifican para las enzimas de la vía de quinurenina — es decir quinureninasa y ácido 3-hidroxiantranil 3,4-dioxigenasa — conducen a malformaciones congénitas vertebrales, anales, cardíacas, traqueoesofágicas, renales y de las extremidades (VACTERL) combinadas (76). Se encontró que el agotamiento de NAD+, en lugar de la acumulación de metabolitos intermedios en la vía de quinurenina era responsable de estas malformaciones. La suplementación con niacina a lo largo del embarazo aseguró niveles adecuados de NAD+ y previno anomalías congénitas en ratones con mutaciones en la vía de quinurenina (76). En los humanos, la dosis de precursores de NAD+ necesaria para evitar malformaciones congénitas VACTERL inducidas por deficiencia de NAD falta aun por definirse (77).
La nicotinamida puede también rescatar la depleción de NAD+ secundaria a un error innato ultra raro del metabolismo de la glutamina (78). La glutamina es requerida para la conversión del ácido nicotínico adenina dinucleótido a NAD+ catalizada por la NAD+ sintetasa (Figura 2). De este modo, la deficiencia de glutamina sintetasa hereditaria específicamente afecta la síntesis de NAD+ a partir de los precursores de NAD+, triptófano y ácido nicotínico. Si las deficiencias combinadas de glutamina y NAD+ son responsables por el fenotipo clínico severo de sujetos con deficiencia hereditaria de glutamina sintetasa, es probable que la suplementación con ambos glutamina y nicotinamida proporcionaría algo de alivio (78).
Finalmente, muchos errores innatos del metabolismo resultan de mutaciones genéticas que disminuyen la afinidad de unión del cofactor y, subsecuentemente, la eficacia de la enzima (79). En muchos casos, la administración de altas dosis de las vitaminas que sirven como precursores de cofactores puede restaurar la actividad enzimática — por lo menos parcialmente — y disminuir los signos de las enfermedades genéticas (79). Dado que un gran numero de enzimas requieren NAD, se especula que muchas de las condiciones debidas a enzimas defectuosas podrían ser rescatadas por la suplementación de niacina (5).
Enfermedades cardiovasculares
Factores de riesgo cardiovascular
El ácido nicotínico es un agente hipolipemiante bien conocido: la terapia con ácido nicotínico aumenta notablemente las concentraciones de colesterol de lipoproteínas de alta densidad (HDL), disminuyendo las concentraciones séricas de lipoproteína(a) y desplaza partículas de lipoproteínas de baja densidad (LDL) pequeñas y densas a partículas LDL grandes y boyantes. Todos estos cambios en el perfil sanguíneo lipídico son considerados cardioprotectores. Bajas concentraciones de colesterol HDL son un factor de gran riesgo para la enfermedad coronaria cardiaca (ECC), y un incremento en las concentraciones HDL esta asociado con una reducción de ese riesgo (80). Debido a los efectos secundarios adversos asociados con las altas dosis de ácido nicotínico (véase Seguridad), el ácido nicotínico ha sido utilizado a menudo en combinación con otros medicamentos hipolipemiantes en dosis ligeramente menores (17). En particular, el ácido nicotínico en dosis bajas es frecuentemente coadministrado con inhibidores de la 3-hidroxi-3-metilglutaril-coenzima A reductasa (estatinas), la piedra angular del tratamiento para la hiperlipidemia, un factor de riesgo importante para la ECC. Un estudio controlado con placebo en 39 pacientes que tomaron estatinas (cerivastatina, atorvastatina o simvastatina) encontró que una dosis muy baja de ácido nicotínico (100 mg/día) incremento el colesterol HDL en solo 2.1 mg/dL, y la combinación no tuvo efecto en las concentraciones de colesterol LDL, colesterol total, o triglicéridos (81).
El estudio de la Biología Arterial para la Investigación de los Efectos del Tratamiento de Reducción del Colesterol (ARBITER) 2 — un ensayo doble ciego, controlado con placebo — investigó el efecto incremental de la adición de niacina (1 g/día) a la terapia con estatinas en 167 pacientes con ECC y niveles bajos de HDL en el grosor íntima-media carotídeo (CIMT) (82), un punto final sustituto para el desarrollo de la aterosclerosis. La adición del ácido nicotínico de liberación prolongada a la simvastatina previene el aumento en CIMT en comparación a la monoterapia de simvastatina. Un análisis post-hoc de datos del ARBITER2 mostró que el bloqueo de la progresión de la aterosclerosis se relacionaba con el aumento en las concentraciones de HDL en pacientes con un estado glucémico normal. Sin embargo, en la presencia de factores de riesgo adicionales, como la glucosa en ayunas alterada o la diabetes mellitus, el incremento de las concentraciones de HDL no fue predictivo de reducción del CIMT y retraso aterosclerótico (83). Un ensayo comparativo de eficacia (ARBITER6) también mostro una reducción significativa del CIMT basal con ácido nicotínico de liberación extendida (2 g/día por 14 meses), a diferencia de ezetimibe (un fármaco hipolipemiante) en pacientes que tomaban estatinas (84).
Estudios adicionales han examinado el impacto del acido nicotínico en la dilatación mediada por flujo (DMF) braquial dependiente del endotelio en pacientes en riesgo de ECC o con ECC establecida. La medición de la DMF es frecuentemente usada como un marcador sustituto de la función endotelial; los valores de la DMF están inversamente correlacionados con el riesgo de eventos cardiovasculares futuros (85). Un meta-análisis de siete ensayos controlados aleatorios, que incluyo 441 participantes, mostro un incremento significante del 2% con el acido nicotínico (1-2 g/día) administrado por 12 semanas a un año (86).
Eventos cardiovasculares
Varios ensayos aleatorios, controlados con placebo, multicéntricos han investigado la eficacia y seguridad de la terapia con ácido nicotínico, sola o en combinación con otros agentes hipolipemiantes en los resultados de las enfermedades cardiovasculares (ECV). Específicamente, el Proyecto de Drogas Coronarias (CDP) dio seguimiento a mas de 8,000 hombres con un infarto al corazón previo por seis años (87). En comparación al grupo de placebo, los pacientes que tomaron diariamente 3 g de ácido nicotínico de liberación inmediata una reducción del 10% en el colesterol total, una disminución del 26% en los triglicéridos, una reducción del 27% en infartos al miocardio no letales recurrentes, y una reducción del 26% en eventos cerebrovasculares (accidente cerebrovascular, y ataque isquémico transitorio). Además, un ensayo posterior de nueve años de seguimiento reveló una reducción del 10% en el total de muertes con el tratamiento con ácido nicotínico.
El Estudio del Tratamiento de la Aterosclerosis HDL (HATS), un ensayo controlado aleatorio de tres años en 160 pacientes con ECC documentada y concentraciones bajas de HDL, encontró que una combinación de simvastatina y ácido nicotínico (2-3 g/día) incrementó los niveles HDL, inhibió la progresión de la estenosis (estrechamiento) de la arteria coronaria, y disminuyó la frecuencia de eventos cardiovasculares, incluyendo el infarto al miocardio y accidentes cerebrovasculares, en comparación al placebo (88). Un análisis de subgrupo de pacientes del HATS con síndrome metabólico mostro una reducción en la tasa de eventos clínicos primarios, aunque la glucosa y el metabolismo de la insulina estaban moderadamente alterados por el acido nicotínico (89). Además, una revisión de la seguridad y tolerabilidad al ácido nicotínico entre los sujetos del HATS mostró que el control glucémico en pacientes con diabetes volvió a los valores de pretratamiento luego de ocho meses del manejo de la enfermedad con medicamento y dieta (90). Similarmente, el beneficio cardiovascular de la terapia con acido nicotínico a largo plazo superó el modesto aumento en el riesgo de una nueva aparición de diabetes tipo 2 en pacientes del estudio CDP (91).
En contraste, el ensayo AIM-HIGH (Intervención de la aterotrombosis en el síndrome metabólico con bajo HDL/triglicéridos altos: impacto en los resultados salud global), el cual examino el efecto incremental del ácido nicotínico de liberación extendida (1.5-2 g/día) en 3,414 pacientes que tenían ECV establecidas y dislipidemia aterogénica y eran tratados con simvastatina (+/- ezetimibe), proporcionó resultados decepcionantes. Ciertamente, en estos pacientes que habían alcanzado concentraciones objetivo de colesterol LDL (<70 mg/dL) antes de la aleatorización, el efecto de elevación de HDL del tratamiento con ácido nicotínico falló en reducir el numero de eventos cardiovasculares después de un seguimiento promedio de tres años (92, 93). Mientras que algunas de las limitaciones, como un uso mayor de simvastatina y ezetimibe en el grupo de control, han confundido los resultados, se sugirió también que el colesterol HDL bajo podría ser un marcador del riesgo en lugar de ser un factor de riesgo causal para la predicción de ECV (93). Además, un análisis post-hoc de 505 participantes con enfermedad crónica renal en etapa 3 encontró un incremento en la mortalidad por todas las causas en aquellos aleatorizados para el ácido nicotínico en comparación a aquellos en el grupo de placebo (94).
Aunque el acido nicotínico falló en reducir el numero de eventos cardiovasculares en pacientes con colesterol LDL bajo tratados con simvastatina, estos resultados no pueden extrapolar a pacientes con colesterol LDL más alto al inicio del estudio. Un ensayo multicéntrico, aleatorio, doble ciego, controlado con placebo de mayor magnitud — el ensayo HPS2-THRIVE (Estudio para la protección del corazón 2: tratamiento de HDL para reducir la incidencia de eventos vasculares) — en 25,673 participantes con enfermedad vascular examinó el efecto incremental del ácido nicotínico de liberación extendida (2 g/día) y el laropiprant (un antagonista del receptor-1 de la prostaglandina D2; 40 mg/día) en la incidencia de eventos cardiovasculares. En comparación al placebo, el ácido nicotínico/laropiprant redujo el colesterol LDL en un promedio de 10 mg/dL, disminuyó los triglicéridos 33 mg/dL, e incrementó el colesterol HDL 6 mg/dL después de un periodo medio de seguimiento de 3.9 años. No obstante, el ácido nicotínico/laropiprant mostró no efecto en la incidencia de eventos vasculares mayores y muerte por cualquier causa (95).
Un reciente meta-análisis de 23 ensayos controlados aleatorios — incluyendo los ensayos CDP, AIM-HIGH y HPS2-THRIVE — en 39,195 sujetos con un historial de enfermedad vascular comparó el efecto del acido nicotínico solo o como un agregado a otros agentes hipolipemiantes. Ningún beneficio cardiovascular fue asociado con la terapia con ácido nicotínico: el numero de infartos al miocardio letales y no letales y accidentes cerebrovasculares no disminuyo con la suplementación de acido nicotínico (dosis mediana de 2 g/día por un periodo medio de 11.5 meses) (96).
A pesar de la falta de evidencia para un papel del acido nicotínico en la prevención de ECV (96, 97), el uso de la terapia con ácido nicotínico ha rápidamente incrementado a través de los años en los Estados Unidos (98).
Ataxia de Friedreich
La ataxia de Friedreich, una forma común de ataxia hereditaria es un trastorno recesivo de aparición temprana con características clínicas que incluyen ataxia progresiva, escoliosis, cardiomiopatía, y diabetes mellitus (99). La mayoría de los sujetos afectados tienen expansiones repetidas homocigotas de guanina-adenina-adenina (GAA) en el primer intrón del gen FXN que codifica para la proteína frataxina. Estas repeticiones GAA anormales e inestables desencadenan el silenciamiento genético a través de la formación de heterocromatina, llevando a una expresión de frataxina significantemente reducida (100). La frataxina es una proteína mitocondrial necesaria para la creación de agrupaciones de hierro y sulfuro (ISC). Las subunidades que contienen ISC son especialmente importantes para la cadena respiratoria mitocondrial y para la síntesis de proteínas que contienen hemo (99).
Predominantemente localizada en el núcleo, SIRT1 es una deacetilasa dependiente de NAD+ que promueve el silenciamiento genético a través de la formación de la heterocromatina. Se ha mostrado que la nicotinamida antagoniza la heterocromatización del locus FXN y regula positivamente la expresión de frataxina en células linfoblastoides derivadas de pacientes afectados por la ataxia de Friedreich, posiblemente a través la inhibición de la actividad de SIRT1 (100). En un ensayo piloto, abierto de aumento de dosis en 10 pacientes adultos con ataxia de Friedreich, encontró que dosis solas y repetidas de nicotinamida (2-8 g) por un periodo de hasta ocho semanas eran bien toleradas (101). Dosis repetidas de 3.5 a 6 g de nicotinamida condujeron a incrementos significantes en la concentración de frataxina en glóbulos blancos periféricos (101). Sin embargo, ninguna mejora neurológica fue reportada, sugiriendo que la duración del tratamiento fue muy corta y/o el sistema nervioso de los participantes no respondió a los aumentos de la frataxina (102). Que sepamos, no existe actualmente algún ensayo en progreso diseñado para investigar más allá el efecto de la nicotinamida en pacientes afectados con la ataxia de Friedreich.
VIH/SIDA
El primer paso en la vía de quinurenina es catalizado por la enzima extrahepática, indoleamina 2,3-dioxigenasa (IDO), la cual es responsable por la escisión oxidativa del triptófano. La estimulación crónica de la oxidación del triptófano, mediada por un incremento en la actividad de IDO y/o las ingestas dietarías inadecuadas de niacina, es observada por la infección del virus de inmunodeficiencia humana (VIH), el virus que causa el síndrome de inmunodeficiencia adquirida (SIDA). El interferón-gamma (IFN- γ) es una citoquina producida por células del sistema inmunitario en respuesta a la infección. A través de la estimulación de la enzima IDO, el IFN- γ incrementa la descomposición del triptófano, apoyando de esta manera el descubrimiento de que la concentración de triptófano promedio en la sangre es significantemente menor en pacientes con VIH en comparación con sujetos no infectados (40). Una degradación incrementada del triptófano a través de la vía de quinurenina parece coexistir con la deficiencia intracelular de niacina/NAD en la infección con VIH (103). Un modelo explicativo de estas observaciones paradójicas incrimina al estrés oxidativo inducido por múltiples deficiencias en los pacientes con VIH (103). En particular la activación de enzimas PARP (ARTD) por el daño oxidativo al ADN podría ser responsable por inducir el agotamiento de niacina/NAD (véase Función). La descomposición del triptófano seria entonces una respuesta compensatoria a los niveles de niacina/NAD inadecuados.
Sin embargo, los metabolismos derivados de la oxidación del triptófano en la vía de quinurenina regulan subgrupos de linfocitos T específicos. Como fue mencionado anteriormente, el IFN-γ circulante, pero también los productos virales y bacterianos, pueden activar la IDO durante la infección con VIH. La sobreestimulación de la ruta del triptófano ha sido involucrada en la perdida de la función normal de los linfocitos T, que caracteriza la infección por VIH (104, 105). El aumento en la actividad de la IDO ha sido ligado a la respuesta inmune alterada que contribuye a la persistencia del VIH (104). La terapia antirretroviral (TARV) solo restaura parcialmente la actividad normal de la IDO, sin normalizarla, pero induce la supresión viral y la recuperación de células T CD4 (106). En un modelo de mono para la infección por VIH, un bloqueo parcial y transitorio de la IDO con el inhibidor de IDO 1-metil-triptófano probó ser ineficaz para reducir la carga viral en el plasma y los tejidos intestinales más allá de los niveles logrados por la TARV (107). En la actualidad, un mejor entendimiento del papel de la vía de quinurenina y otras rutas NAD biosintéticas durante la infección por VIH es necesario antes de que se pueda considerar la relevancia y las implicaciones clínicas de la administración de suplementos de niacina en el tratamiento del VIH.
No obstante, las dosis farmacológicas de ácido nicotínico han mostrado ser bien toleradas en los pacientes con VIH con hiperlipidemia (108). Los perfiles de lípidos anormales observados en pacientes han sido atribuidos a la infección por VIH y al tratamiento antirretroviral altamente activo (TARAA) (109). Además, la resistencia a la insulina ha sido detectada junto con dislipidemia en pacientes tratados con TARV (110). Las enfermedades cardiovasculares (ECV) son la segunda causa más frecuente de muerte en la población con VIH, y se predice que la tasa de ECV incrementara aún más ya que los pacientes están viviendo más debido a las terapias antirretrovirales exitosas. Con respecto a la población general, la terapia basada en estatinas parece beneficiar los pacientes con VIH en términos de protección aterogénica y reducción del riesgo de ECV, aunque contraindicaciones existen debido a la interacción de drogas con la TARV. Otros tratamientos de primera línea incluyen fibratos hipolipemiantes, los cuales son preferidos al acido nicotínico debido al riesgo incrementado de intolerancia a la glucosa y resistencia a la insulina (111). Sin embargo, un estudio piloto controlado, sin cegar mostró que el ácido nicotínico de liberación extendida (0.5-1.5 g/día por 12 semanas) pudo efectivamente mejorar la función endotelial de la arteria braquial en los sujetos con VIH tratados con TARV con bajo colesterol HDL y sin historial de ECV (112). Además, un tratamiento combinado de fibratos, acido nicotínico de liberación extendida (0.5-2 g/día), y cambios en el estilo de vida (dieta baja en grasa y ejercicio) por 24 semanas fue eficaz en la normalización de los parámetros lipídicos en una cohorte de 191 pacientes tratados con TARV. El riesgo incrementado de disfunción hepática fue detectado en sujetos que recibían ambos fibratos y niacina, pero la sensibilidad a la insulina no fue afectada por el tratamiento con ácido nicotínico administrado solo o cuando fue combinado con fibratos (113). Otro ensayo no controlado, abierto, de 24 semanas en 99 pacientes tratados con TARV encontró que la aleatorización de ácido nicotínico de liberación extendida (0.5-2 g/día) o fenofibratos incrementó el colesterol HDL sanguíneo, pero no redujo los marcadores inflamatorios o mejoro la función endotelial en comparación al valor basal (114).
Esquizofrenia
La esquizofrenia es un trastorno neurológico de etiología poco clara que se diagnostica solo a partir de su presentación clínica. Debido a que los trastornos neurológicos asociados con la pelagra se asemejan a la esquizofrenia aguda, la terapia basada en niacina para la condición fue investigada durante 1950-70 (revisado en 115). El uso adyuvante de nutrientes como la niacina para corregir las deficiencias asociadas con síntomas neurológicos se llama psiquiatría ortomolecular (116). Tal enfoque no se ha incluido en la práctica psiquiátrica; los profesionales se han basado únicamente en fármacos antipsicóticos para eliminar los síntomas clínicos de la esquizofrenia. Sin embargo, avances científicos recientes y nuevas hipótesis en el beneficio de la suplementación de nutrientes en el tratamiento de trastornos psiquiátricos han sugerido la reevaluación de la medicina ortomolecular por la comunidad médica (117, 118).
El enrojecimiento de la piel es uno de los principales efectos secundarios del uso terapéutico del ácido nicotínico y la razón principal de la falta de adherencia al tratamiento (ver Toxicidad). El enrojecimiento es causado por la activación de la fosfolipasa A2, una enzima que estimula la producción de un lípido específico de la familia prostanoide llamado prostaglandina D2. La prostaglandina D2, sintetizada por las células presentadoras de antígenos de la piel y la mucosa (es decir, las células de Langerhans), puede inducir la dilatación de los vasos sanguíneos y desencadenar una respuesta de enrojecimiento. Curiosamente, los pacientes con esquizofrenia tienden a no enrojecer luego del tratamiento con ácido nicotínico. Esta respuesta de enrojecimiento de piel deficiente sugiere una señalización prostanoide en pacientes esquizofrénicos (119, 120). Se ha encontrado una asociación entre la sensibilidad de niacina alterada y un mayor deterioro funcional en pacientes esquizofrénicos (121), que apoya otros hallazgos que sugieren que el metabolismo lipídico alterado podría críticamente alterar el desarrollo del cerebro y contribuir a la enfermedad (122). Curiosamente, las respuestas de enrojecimiento cutáneas deficientes son más prevalentes en familiares de primer grado de personas con esquizofrenia que en la población general, sugiriendo que la sensibilidad reducida a la niacina es un rasgo hereditario dentro de las familias afectadas (123).
Fuentes
Fuentes alimenticias
Buenas fuentes de niacina incluyen la levadura, carne, aves de corral, pescados rojos (p.ej. atún, salmón), los cereales (especialmente los cereales fortificados), legumbres, y semillas. La leche, verduras de hojas verdes, el café y té también aportan una pequeña cantidad de niacina (124). En las plantas, especialmente los granos de cereal maduro como el maíz y el trigo, la niacina podría estar ligada a moléculas de azúcar en la forma de glucósidos, lo que disminuye significativamente su biodisponibilidad (25).
En los Estados Unidos, la ingesta dietaría promedio de niacina es de alrededor de 30 mg/día para hombres adultos jóvenes y de 20 mg/día para las mujeres adultas jóvenes. En una muestra de adultos mayores de 60 años, se encontró que hombres y mujeres tenían una ingesta dietaría promedio de 21 mg/día y 17 mg/día, respectivamente (41). Algunos alimentos con cantidades sustanciales de niacina son listados en la Tabla 2, junto con su contenido de niacina en miligramos (mg). Las tablas de composición de los alimentos generalmente muestran el contenido de niacina sin incluir los equivalentes de niacina (EN) del triptófano o algún ajuste para la biodisponibilidad de niacina. Para mayor información en el contenido de nutrientes de alimentos específicos, revise la base de datos de composición de los alimentos del USDA; la información incluida en la Tabla 2 proviene de esta base de datos (125).
Suplementos
Los suplementos de niacina están disponibles como nicotinamida o acido nicotínico. La nicotinamida es la forma de la niacina típicamente usada en suplementos nutricionales y en la fortificación de alimentos. El ácido nicotínico esta disponible sin receta y con una prescripción como un agente reductor del colesterol (126). El ácido nicotínico para su uso anti-hiperlipidémico en tres formulaciones: acido nicotínico de liberación inmediata (cristalino) (tiempo de absorción, 1-2 horas), acido nicotínico de liberación extendida (tiempo de absorción, 8-12 horas), y acido nicotínico de liberación sostenida (tiempo de absorción, >12 horas) (127). En la dosis farmacológica requerida para los efectos de reducción del colesterol, el uso de ácido nicotínico debe ser abordado como si este fuese un fármaco (véase Seguridad). Las personas solo deben tomar una terapia de reducción del colesterol con ácido nicotínico bajo la supervisión de un proveedor del cuidado de la salud calificado con el fin de minimizar los efectos potencialmente adversos y maximizar los beneficios terapéuticos.
Seguridad
Toxicidad
A la niacina de los alimentos no se le conoce por causar efectos adversos. Aunque un estudio notó efectos adversos luego del consumo de bagels con 60 veces más la cantidad normal de la fortificación con niacina, la mayoría de los efectos han sido reportados con preparaciones farmacológicas de niacina (41).
Acido nicotínico
Los efectos secundarios comunes de ácido nicotínico incluyen enrojecimiento, prurito (picazón severa de la piel), erupciones cutáneas y trastornos gastrointestinales, como las náuseas y vómitos (97). También se han informado episodios transitorios de presión arterial baja (hipotensión) y dolor de cabeza. La hepatotoxicidad (daño celular hepático), que incluye enzimas hepáticas elevadas e ictericia, ha sido observada en ingestas tan bajas de hasta 750 mg/día de ácido nicotínico (128). Aunque la hepatitis ha sido observada con dosis de ácido nicotínico de liberación extendida de tan poco como 500 mg/día por dos meses, casi todos los reportes de hepatitis severa han sido asociados con dosis de 3 a 9 g/día usados para tratar el colesterol alto por meses o años (41). No esta claro si el ácido nicotínico de liberación inmediata (cristalino) es menos toxico para el hígado que las formas de liberación extendida (41). Sin embargo, el ácido nicotínico de liberación inmediata es frecuentemente usado en dosis más altas que las formas de liberación extendida, y la toxicidad severa hepática ha ocurrido en individuos que sustituyeron el ácido nicotínico de liberación extendida por acido nicotínico de liberación inmediata en dosis equivalentes (126). Se ha observado que grades dosis de ácido nicotínico alteran la tolerancia a la glucosa probablemente debido a una disminución en la sensibilidad a la insulina. La tolerancia a la glucosa alterada en individuos susceptibles (prediabéticos) podría resultar en concentraciones elevadas de glucosa en la sangre y diabetes mellitus tipo 2. Un análisis del ensayo HPS2-THRIVE (véase Enfermedades cardiovasculares), usando datos de 17,374 participantes sin diabetes tipo 2 al inicio del estudio, encontró una proporción significantemente mas alta de casos recientemente diagnosticados entre aquellos aleatorizados para el ácido nicotínico/laropiprant que para el placebo (5.7% versus 4.3%) sobre un periodo de 3.9 años (95). De igual manera, la aleatorización para el ácido nicotínico/laropiprant significantemente incremento el riesgo de disturbios serios en el control para la diabetes (llevando a la hospitalización) en comparación al placebo entre 8,299 participantes con diabetes al inicio del estudio (95). Las concentraciones sanguíneas elevadas de ácido úrico, ocasionalmente resultando en ataques de gota en los individuos susceptibles, han sido también observados con la terapia con ácido nicotínico de alta dosis (126). La niacina en dosis de 1.5 a 5 g/día ha resultado en unos pocos reportes de caso de visión borrosa y otros problemas oculares, los cuales han sido generalmente reversibles luego de la discontinuación (41). Personas con una función hepática anormal o un historial de enfermedad hepática, diabetes, enfermedad de úlcera péptica activa, gota, arritmias cardiacas, enfermedad intestinal inflamatoria, migrañas, o alcoholismo pueden ser mas susceptibles a los efectos adversos de la ingesta de niacina en exceso que la población en general (41).
Nicotinamida
La nicotinamida es generalmente mejor tolerada que el acido nicotínico; generalmente no causa enrojecimiento (126). Sin embargo, náuseas, vómitos, y signos de toxicidad hepática (enzimas hepáticas elevadas, ictericia) han sido observados en dosis muy altas (≥10 g/día) (126).
Nicotinamida ribósido
Un estudio en 12 sujetos saludables encontró que la nicotinamida ribósido en tres dosis únicas (100 mg, 300 mg, y 1,000 mg) incremento el NAD+ sanguíneo sin peligro. Dos de los participantes auto-reportaron enrojecimiento de la piel después de tomar la dosis de 300 mg, y dos otros reportaron una sensación de calor luego de la ingesta de 1,000 mg de nicotinamida ribósido (1). En un ensayo aleatorio, controlado con placebo reciente en 120 adultos saludables (edades, 60-80 años), la suplementación diaria con nicotinamida ribósido (250 mg o 500 mg) y pteroestilbeno (un activador de SIRT; 50 mg o 100 mg) por ocho semanas mostro un perfil favorable de efectos secundarios, sin evidencia de una incidencia mayor de efectos adversos en comparación al placebo (129). Más recientemente, un ensayo aleatorio, controlado con placebo en 40 hombres obesos (edades, 40-70 años) encontró que la suplementación diaria con nicotinamida ribósido (2,000 mg/día dividida en dos dosis diarias) durante 12 semanas fue asociada con reportes de solo efectos secundarios, incluyendo sudoración excesiva, prurito y síntomas gastrointestinales leves como la hinchazón (139).
El nivel máximo de ingesta tolerable (NM)
El enrojecimiento de la piel principalmente en la cara, brazos, y pecho, es un efecto secundario común del ácido nicotínico y puede ocurrir inicialmente con dosis tan bajas como de 30 mg/día. Aunque el enrojecimiento causado por la nicotinamida es raro, la Junta de Nutrición y Alimentos estableció el nivel máximo de ingesta tolerable (NM) para la niacina (ácido nicotínico y nicotinamida) en 35 mg/día para los adultos para evitar el enrojecimiento como un efecto adverso (41). El análisis de datos de la Encuesta de Evaluación Nacional de Salud y Nutrición Estadounidense (NHANES) 2003-2006 encontró que el 15.8% de los niños y adolescentes (edades 2-18 años) y el 8.5% de los adultos (≥19 años) tuvieron ingestas usuales de niacina totales que excedían el NM (130). El NM se aplica a toda la población en general y no esta destinado a ser aplicado en individuos que están siendo tratados con un nutriente bajo supervisión médica (p. ej., acido nicotínico en altas dosis para las concentraciones elevadas de colesterol sanguíneo).
| Grupo Etario | NM (mg/día) |
|---|---|
| Infantes 0-12 meses | Imposible de determinar* |
| Niños 1-3 años | 10 |
| Niños 4-8 años | 15 |
| Niños 9-13 años | 20 |
| Adolescentes 14-18 años | 30 |
| Adultos 19 años y más | 35 |
| *La fuente de la ingesta debiera ser sólo de alimentos y fórmula. | |
Interacción con drogas/fármacos
La ocurrencia de rabdomiolisis se incrementa en los pacientes tratados con estatinas (inhibidores de la HMG-CoA reductasa). La rabdomiolisis es una condición relativamente poco común en la cual las células musculares se degradan, liberando enzimas y electrolitos a la sangre, y algunas veces resultando en falla renal (131). La coadministración de ácido nicotínico con una estatina parece incrementar el riesgo de rabdomiolisis (132). Un nuevo fármaco, laropiprant, bloquea los receptores prostanoides y reduce el enrojecimiento inducido por el ácido nicotínico (133). Un ensayo aleatorio, controlado con placebo fue designado para identificar efectos adversos posibles de la combinación de niacina/laropiprant en más de 25,000 pacientes tratados con simvastatina (134). Cuando se agregó a la terapia con estatinas, la combinación de niacina/laropiprant aumentó el riesgo de miopatía y rabdomiolisis, particularmente en sujetos asiáticos. Es posible que la combinación de niacina/laropirant reduzca aún más la baja tolerabilidad del tratamiento con estatinas observadas en ciertas poblaciones (135).
En el estudio aleatorio controlado HATS de tres años, la terapia concurrente con antioxidantes (1,000 mg/día de vitamina C, 800 UI/día de RRR-α-tocoferol, 100 µg/día de selenio, y 25 mg/día de β-caroteno) disminuyó los efectos protectores de la combinación de simvastatina-acido nicotínico (136). Aunque el mecanismo para estos efectos es desconocido, el beneficio de la terapia antioxidante concurrente en pacientes que toman agentes hipolipemiantes ha sido cuestionado (137).
Los efectos adversos de grandes dosis de ácido nicotínico pueden ser exacerbados por el uso concomitante de ciertos medicamentos. El riesgo de miopatía puede aumentar aún más en aquellos que toman ácido nicotínico y secuestrantes de ácidos biliares (p. ej., colestiramina, colestipol) o la droga antilipidémica, gemfibrozil (Lopid), y el riesgo de hepatotoxicidad observado con el ácido nicotínico podría ser incrementado por fármacos como el paracetamol, la amiodarona (Cordarone) o la carbamazepina (Tegretol) (35). Además, grandes dosis de ácido nicotínico pueden reducir la excreción de ácido úrico, oponiéndose de este modo a la acción de los agentes uricosúricos como el probenecid (Probalan) (35).
Varios otros medicamentos pueden interactuar con la terapia con niacina o con la absorción y metabolismo de la vitamina (126). El estrógeno y los anticonceptivos que contienen estrógeno incrementan la eficacia de la síntesis de niacina a partir del triptófano, resultando en un requerimiento dietario disminuido para la niacina (138).
Se ha reportado que la administración a largo plazo de agentes de la quimioterapia causa síntomas de pelagra; por lo tanto, la suplementación con niacina puede ser necesaria (véase Causas de la pelagra).
Recomendación del Instituto Linus Pauling
La ingesta optima de niacina para la promoción de la salud y prevención de enfermedades crónicas no es aun conocida. La IDR (16 mg EN/día para hombres y 14 mg EN/día para mujeres) es obtenible fácilmente consumiendo una dieta variada y debería prevenir una deficiencia en la mayoría de las personas. Siguiendo la recomendación del Instituto Linus Pauling de tomar a diario un suplemento multivitamínico/mineral que contenga el 100% del Valor Diario (VD) para niacina, aportará diariamente al menos 20 mg de niacina.
Autores y Críticos
Escrito en 2002 por:
Jane Higdon, Ph.D.
Instituto Linus Pauling
Universidad Estatal de Oregon
Actualizado en Agosto 2002 por:
Jane Higdon, Ph.D.
Instituto Linus Pauling
Universidad Estatal de Oregon
Actualizado en Junio 2007 por:
Victoria J. Drake, Ph.D
Instituto Linus Pauling
Universidad Estatal de Oregon
Actualizado en Julio 2013 por:
Barbara Delage, Ph.D.
Instituto Linus Pauling
Universidad Estatal de Oregon
Actualizado en Diciembre de 2017 por:
Barbara Delage, Ph.D.
Instituto Linus Pauling
Universidad Estatal de Oregon
Revisado en Marzo de 2018 por:
Mirella Meyer-Ficca, Ph.D.
Profesor Asistente de Investigación
Universidad del Estado de Utah
Traducido al Español en 2018 por:
Silvia Vazquez Lima
Instituto Linus Pauling
Universidad Estatal de Oregon
La actualización y traducciones al español de este artículo en el 2017 fue respaldada por una subvención de ChromaDex, Inc.
Originalmente traducido al español en 2012 por Guillermo Sandoval y editado por Andrew Quest (Ph.D.) y Lisette Leyton (Ph.D.), todos provenientes de la Universidad de Chile. Estos esfuerzos fueron patrocinados por el projecto Anillo #ACT1111, CONICYT-Chile, programa PIA.
Derechos de autoría 2000-2024 Instituto Linus Pauling
Referencias
1. Trammell SA, Schmidt MS, Weidemann BJ, et al. Nicotinamide riboside is uniquely and orally bioavailable in mice and humans. Nat Commun. 2016;7:12948. (PubMed)
2. Nikiforov A, Kulikova V, Ziegler M. The human NAD metabolome: functions, metabolism and compartmentalization. Crit Rev Biochem Mol Biol. 2015;50(4):284-297. (PubMed)
3. Kawai S, Murata K. Structure and function of NAD kinase and NADP phosphatase: key enzymes that regulate the intracellular balance of NAD(H) and NADP(H). Biosci Biotechnol Biochem. 2008;72(4):919-930. (PubMed)
4. Agledal L, Niere M, Ziegler M. The phosphate makes a difference: cellular functions of NADP. Redox Rep. 2010;15(1):2-10. (PubMed)
5. Penberthy WT, Kirkland JB. Niacin. In: Erdman JW, MacDonald I, Zeisel SH, eds. Present Knowledge in Nutrition. 10th ed. Ames: International Life Sciences Institute; 2012:293-306.
6. Kirkland JB. Niacin. In: Ross AC, Caballero B, Cousins RJ, Tucker KL, Ziegler TR, eds. Modern Nutrition in Health and Disease. 11th ed. Baltimore: Lippincott Williams & Wilkins; 2014:331-340.
7. Hottiger MO, Hassa PO, Luscher B, Schuler H, Koch-Nolte F. Toward a unified nomenclature for mammalian ADP-ribosyltransferases. Trends Biochem Sci. 2010;35(4):208-219. (PubMed)
8. Liu C, Yu X. ADP-ribosyltransferases and poly ADP-ribosylation. Curr Protein Pept Sci. 2015;16(6):491-501. (PubMed)
9. Hwang ES, Song SB. Nicotinamide is an inhibitor of SIRT1 in vitro, but can be a stimulator in cells. Cell Mol Life Sci. 2017;74(18):3347-3362. (PubMed)
10. Morris BJ. Seven sirtuins for seven deadly diseases of aging. Free Radic Biol Med. 2013;56:133-171. (PubMed)
11. Fliegert R, Bauche A, Wolf Perez AM, et al. 2'-Deoxyadenosine 5'-diphosphoribose is an endogenous TRPM2 superagonist. Nat Chem Biol. 2017;13(9):1036-1044. (PubMed)
12. Mutafova-Yambolieva VN, Hwang SJ, Hao X, et al. Beta-nicotinamide adenine dinucleotide is an inhibitory neurotransmitter in visceral smooth muscle. Proc Natl Acad Sci U S A. 2007;104(41):16359-16364. (PubMed)
13. Moreschi I, Bruzzone S, Nicholas RA, et al. Extracellular NAD+ is an agonist of the human P2Y11 purinergic receptor in human granulocytes. J Biol Chem. 2006;281(42):31419-31429. (PubMed)
14. Klein C, Grahnert A, Abdelrahman A, Muller CE, Hauschildt S. Extracellular NAD(+) induces a rise in [Ca(2+)](i) in activated human monocytes via engagement of P2Y(1) and P2Y(11) receptors. Cell Calcium. 2009;46(4):263-272. (PubMed)
15. Moreschi I, Bruzzone S, Bodrato N, et al. NAADP+ is an agonist of the human P2Y11 purinergic receptor. Cell Calcium. 2008;43(4):344-355. (PubMed)
16. Huang C, Hu J, Subedi KP, et al. Extracellular adenosine diphosphate ribose mobilizes intracellular Ca2+ via purinergic-dependent Ca2+ pathways in rat pulmonary artery smooth muscle cells. Cell Physiol Biochem. 2015;37(5):2043-2059. (PubMed)
17. Knopp RH. Drug treatment of lipid disorders. N Engl J Med. 1999;341(7):498-511. (PubMed)
18. Graff EC, Fang H, Wanders D, Judd RL. Anti-inflammatory effects of the hydroxycarboxylic acid receptor 2. Metabolism. 2016;65(2):102-113. (PubMed)
19. Jin FY, Kamanna VS, Kashyap ML. Niacin accelerates intracellular ApoB degradation by inhibiting triacylglycerol synthesis in human hepatoblastoma (HepG2) cells. Arterioscler Thromb Vasc Biol. 1999;19(4):1051-1059. (PubMed)
20. Kamanna VS, Ganji SH, Kashyap ML. Recent advances in niacin and lipid metabolism. Curr Opin Lipidol. 2013;24(3):239-245. (PubMed)
21. Carlson LA. Studies on the effect of nicotinic acid on catecholamine stimulated lipolysis in adipose tissue in vitro. Acta Med Scand. 1963;173:719-722. (PubMed)
22. Lauring B, Taggart AK, Tata JR, et al. Niacin lipid efficacy is independent of both the niacin receptor GPR109A and free fatty acid suppression. Sci Transl Med. 2012;4(148):148ra115. (PubMed)
23. Brody T. Nutritional Biochemistry. 2nd ed. San Diego: Academic Press; 1999.
24. Kirkland JB. Niacin. In: Zempleni J, Suttie JW, Gregory III JF, Stover PJ, eds. Handbook of Vitamins. 5th ed. Boca Raton: CRC Press; 2013:149-190.
25. Gregory JF, 3rd. Nutritional properties and significance of vitamin glycosides. Annu Rev Nutr. 1998;18:277-296. (PubMed)
26. Dawson B, Favaloro EJ, Taylor J, Aggarwal A. Unrecognized pellagra masquerading as odynophagia. Intern Med J. 2006;36(7):472-474. (PubMed)
27. Jagielska G, Tomaszewicz-Libudzic EC, Brzozowska A. Pellagra: a rare complication of anorexia nervosa. Eur Child Adolesc Psychiatry. 2007;16(7):417-420. (PubMed)
28. Kertesz SG. Pellagra in 2 homeless men. Mayo Clin Proc. 2001;76(3):315-318. (PubMed)
29. Prakash R, Gandotra S, Singh LK, Das B, Lakra A. Rapid resolution of delusional parasitosis in pellagra with niacin augmentation therapy. Gen Hosp Psychiatry. 2008;30(6):581-584. (PubMed)
30. Badawy AA. Pellagra and alcoholism: a biochemical perspective. Alcohol Alcohol. 2014;49(3):238-250. (PubMed)
31. Majewski M, Kozlowska A, Thoene M, Lepiarczyk E, Grzegorzewski WJ. Overview of the role of vitamins and minerals on the kynurenine pathway in health and disease. J Physiol Pharmacol. 2016;67(1):3-19. (PubMed)
32. Rosmaninho A, Sanches M, Fernandes IC, et al. Letter: Pellagra as the initial presentation of Crohn disease. Dermatol Online J. 2012;18(4):12. (PubMed)
33. Zaraa I, Belghith I, El Euch D, et al. A case of pellagra associated with megaduodenum in a young woman. Nutr Clin Pract. 2013;28(2):218-222. (PubMed)
34. Bilgili SG, Karadag AS, Calka O, Altun F. Isoniazid-induced pellagra. Cutan Ocul Toxicol. 2011;30(4):317-319. (PubMed)
35. Natural Medicines. Professional Monograph - Niacin/Interactions with drugs. Available at: https://naturalmedicines.therapeuticresearch.com/. Accessed 8/2/17.
36. Dreizen S, McCredie KB, Keating MJ, Andersson BS. Nutritional deficiencies in patients receiving cancer chemotherapy. Postgrad Med. 1990;87(1):163-167, 170. (PubMed)
37. Nogueira A, Duarte AF, Magina S, Azevedo F. Pellagra associated with esophageal carcinoma and alcoholism. Dermatol Online J. 2009;15(5):8. (PubMed)
38. Oldham MA, Ivkovic A. Pellagrous encephalopathy presenting as alcohol withdrawal delirium: a case series and literature review. Addict Sci Clin Pract. 2012;7(1):12. (PubMed)
39. World Health Organization, United Nations High Commissions for Refugees. Pellagra and its prevention and control in major emergencies. World Health Organization. 2000. Available at: http://www.who.int/nutrition/publications/emergencies/WHO_NHD_00.10/en/. Accessed 6/20/13.
40. Murray MF. Tryptophan depletion and HIV infection: a metabolic link to pathogenesis. Lancet Infect Dis. 2003;3(10):644-652. (PubMed)
41. Food and Nutrition Board, Institute of Medicine. Niacin. Dietary Reference Intakes: Thiamin, Riboflavin, Niacin, Vitamin B-6, Vitamin B-12, Pantothenic Acid, Biotin, and Choline. Washington, D.C.: The National Academies Press; 1998:123-149. (The National Academies Press)
42. Negrini S, Gorgoulis VG, Halazonetis TD. Genomic instability--an evolving hallmark of cancer. Nat Rev Mol Cell Biol. 2010;11(3):220-228. (PubMed)
43. Kirkland JB. Niacin requirements for genomic stability. Mutat Res. 2012;733(1-2):14-20. (PubMed)
44. Burkle A. Poly(ADP-ribose). The most elaborate metabolite of NAD+. FEBS J. 2005;272(18):4576-4589. (PubMed)
45. Jacobson EL, Shieh WM, Huang AC. Mapping the role of NAD metabolism in prevention and treatment of carcinogenesis. Mol Cell Biochem. 1999;193(1-2):69-74. (PubMed)
46. Spronck JC, Nickerson JL, Kirkland JB. Niacin deficiency alters p53 expression and impairs etoposide-induced cell cycle arrest and apoptosis in rat bone marrow cells. Nutr Cancer. 2007;57(1):88-99. (PubMed)
47. Spronck JC, Kirkland JB. Niacin deficiency increases spontaneous and etoposide-induced chromosomal instability in rat bone marrow cells in vivo. Mutat Res. 2002;508(1-2):83-97. (PubMed)
48. Kostecki LM, Thomas M, Linford G, et al. Niacin deficiency delays DNA excision repair and increases spontaneous and nitrosourea-induced chromosomal instability in rat bone marrow. Mutat Res. 2007;625(1-2):50-61. (PubMed)
49. Dantzer F, Santoro R. The expanding role of PARPs in the establishment and maintenance of heterochromatin. FEBS J. 2013;280(15):3508-3518. (PubMed)
50. El Ramy R, Magroun N, Messadecq N, et al. Functional interplay between Parp-1 and SirT1 in genome integrity and chromatin-based processes. Cell Mol Life Sci. 2009;66(19):3219-3234. (PubMed)
51. Boyonoski AC, Spronck JC, Gallacher LM, et al. Niacin deficiency decreases bone marrow poly(ADP-ribose) and the latency of ethylnitrosourea-induced carcinogenesis in rats. J Nutr. 2002;132(1):108-114. (PubMed)
52. Boyonoski AC, Spronck JC, Jacobs RM, Shah GM, Poirier GG, Kirkland JB. Pharmacological intakes of niacin increase bone marrow poly(ADP-ribose) and the latency of ethylnitrosourea-induced carcinogenesis in rats. J Nutr. 2002;132(1):115-120. (PubMed)
53. Weitberg AB. Effect of nicotinic acid supplementation in vivo on oxygen radical-induced genetic damage in human lymphocytes. Mutat Res. 1989;216(4):197-201. (PubMed)
54. Hageman GJ, Stierum RH, van Herwijnen MH, van der Veer MS, Kleinjans JC. Nicotinic acid supplementation: effects on niacin status, cytogenetic damage, and poly(ADP-ribosylation) in lymphocytes of smokers. Nutr Cancer. 1998;32(2):113-120. (PubMed)
55. Yong LC, Petersen MR. High dietary niacin intake is associated with decreased chromosome translocation frequency in airline pilots. Br J Nutr. 2011;105(4):496-505. (PubMed)
56. Weidele K, Beneke S, Burkle A. The NAD+ precursor nicotinic acid improves genomic integrity in human peripheral blood mononuclear cells after X-irradiation. DNA Repair (Amst). 2017;52:12-23. (PubMed)
57. Jacobson EL. Niacin deficiency and cancer in women. J Am Coll Nutr. 1993;12(4):412-416. (PubMed)
58. Negri E, Franceschi S, Bosetti C, et al. Selected micronutrients and oral and pharyngeal cancer. Int J Cancer. 2000;86(1):122-127. (PubMed)
59. Franceschi S, Bidoli E, Negri E, et al. Role of macronutrients, vitamins and minerals in the aetiology of squamous-cell carcinoma of the oesophagus. Int J Cancer. 2000;86(5):626-631. (PubMed)
60. Gensler HL, Williams T, Huang AC, Jacobson EL. Oral niacin prevents photocarcinogenesis and photoimmunosuppression in mice. Nutr Cancer. 1999;34(1):36-41. (PubMed)
61. Jacobson EL, Kim H, Kim M, et al. A topical lipophilic niacin derivative increases NAD, epidermal differentiation and barrier function in photodamaged skin. Exp Dermatol. 2007;16(6):490-499. (PubMed)
62. Bermudez Y, Benavente CA, Meyer RG, Coyle WR, Jacobson MK, Jacobson EL. Nicotinic acid receptor abnormalities in human skin cancer: implications for a role in epidermal differentiation. PLoS One. 2011;6(5):e20487. (PubMed)
63. Benavente CA, Jacobson EL. Niacin restriction upregulates NADPH oxidase and reactive oxygen species (ROS) in human keratinocytes. Free Radic Biol Med. 2008;44(4):527-537. (PubMed)
64. Benavente CA, Schnell SA, Jacobson EL. Effects of niacin restriction on sirtuin and PARP responses to photodamage in human skin. PLoS One. 2012;7(7):e42276. (PubMed)
65. Park SM, Li T, Wu S, et al. Niacin intake and risk of skin cancer in US women and men. Int J Cancer. 2017;140(9):2023-2031. (PubMed)
66. Chen AC, Martin AJ, Choy B, et al. A phase 3 randomized trial of nicotinamide for skin-cancer chemoprevention. N Engl J Med. 2015;373(17):1618-1626. (PubMed)
67. Minocha R, Damian DL, Halliday GM. Melanoma and nonmelanoma skin cancer chemoprevention: A role for nicotinamide? Photodermatol Photoimmunol Photomed. 2018;34(1):5-12. (PubMed)
68. Orban T, Sosenko JM, Cuthbertson D, et al. Pancreatic islet autoantibodies as predictors of type 1 diabetes in the Diabetes Prevention Trial-Type 1. Diabetes Care. 2009;32(12):2269-2274. (PubMed)
69. Szkudelski T. Streptozotocin-nicotinamide-induced diabetes in the rat. Characteristics of the experimental model. Exp Biol Med (Maywood). 2012;237(5):481-490. (PubMed)
70. Lampeter EF, Klinghammer A, Scherbaum WA, et al. The Deutsche Nicotinamide Intervention Study: an attempt to prevent type 1 diabetes. DENIS Group. Diabetes. 1998;47(6):980-984. (PubMed)
71. Gale EA, Bingley PJ, Emmett CL, Collier T, European Nicotinamide Diabetes Intervention Trial Group. European Nicotinamide Diabetes Intervention Trial (ENDIT): a randomised controlled trial of intervention before the onset of type 1 diabetes. Lancet. 2004;363(9413):925-931. (PubMed)
72. Hedman M, Ludvigsson J, Faresjo MK. Nicotinamide reduces high secretion of IFN-gamma in high-risk relatives even though it does not prevent type 1 diabetes. J Interferon Cytokine Res. 2006;26(4):207-213. (PubMed)
73. Fernandez IC, Del Carmen Camberos M, Passicot GA, Martucci LC, Cresto JC. Children at risk of diabetes type 1. Treatment with acetyl-L-carnitine plus nicotinamide - Case reports. J Pediatr Endocrinol Metab. 2013;26(3-4):347-355. (PubMed)
74. Patel AB, Prabhu AS. Hartnup disease. Indian J Dermatol. 2008;53(1):31-32. (PubMed)
75. Oakley A, Wallace J. Hartnup disease presenting in an adult. Clin Exp Dermatol. 1994;19(5):407-408. (PubMed)
76. Shi H, Enriquez A, Rapadas M, et al. NAD deficiency, congenital malformations, and niacin supplementation. N Engl J Med. 2017;377(6):544-552. (PubMed)
77. Vander Heiden MG. Metabolism and congenital malformations - NAD's effects on development. N Engl J Med. 2017;377(6):509-511. (PubMed)
78. Hu L, Ibrahim K, Stucki M, et al. Secondary NAD+ deficiency in the inherited defect of glutamine synthetase. J Inherit Metab Dis. 2015;38(6):1075-1083. (PubMed)
79. Ames BN, Elson-Schwab I, Silver EA. High-dose vitamin therapy stimulates variant enzymes with decreased coenzyme binding affinity (increased K(m)): relevance to genetic disease and polymorphisms. Am J Clin Nutr. 2002;75(4):616-658. (PubMed)
80. Bays HE, Shah A, Lin J, Sisk CM, Dong Q, Maccubbin D. Consistency of extended-release niacin/laropiprant effects on Lp(a), ApoB, non-HDL-C, Apo A1, and ApoB/ApoA1 ratio across patient subgroups. Am J Cardiovasc Drugs. 2012;12(3):197-206. (PubMed)
81. Wink J, Giacoppe G, King J. Effect of very-low-dose niacin on high-density lipoprotein in patients undergoing long-term statin therapy. Am Heart J. 2002;143(3):514-518. (PubMed)
82. Taylor AJ, Sullenberger LE, Lee HJ, Lee JK, Grace KA. Arterial Biology for the Investigation of the Treatment Effects of Reducing Cholesterol (ARBITER) 2: a double-blind, placebo-controlled study of extended-release niacin on atherosclerosis progression in secondary prevention patients treated with statins. Circulation. 2004;110(23):3512-3517. (PubMed)
83. Taylor AJ, Zhu D, Sullenberger LE, Lee HJ, Lee JK, Grace KA. Relationship between glycemic status and progression of carotid intima-media thickness during treatment with combined statin and extended-release niacin in ARBITER 2. Vasc Health Risk Manag. 2007;3(1):159-164. (PubMed)
84. Villines TC, Stanek EJ, Devine PJ, et al. The ARBITER 6-HALTS Trial (Arterial Biology for the Investigation of the Treatment Effects of Reducing Cholesterol 6-HDL and LDL Treatment Strategies in Atherosclerosis): final results and the impact of medication adherence, dose, and treatment duration. J Am Coll Cardiol. 2010;55(24):2721-2726. (PubMed)
85. Ras RT, Streppel MT, Draijer R, Zock PL. Flow-mediated dilation and cardiovascular risk prediction: a systematic review with meta-analysis. Int J Cardiol. 2013;168(1):344-351. (PubMed)
86. Sahebkar A. Effect of niacin on endothelial function: a systematic review and meta-analysis of randomized controlled trials. Vasc Med. 2014;19(1):54-66. (PubMed)
87. Canner PL, Berge KG, Wenger NK, et al. Fifteen year mortality in Coronary Drug Project patients: long-term benefit with niacin. J Am Coll Cardiol. 1986;8(6):1245-1255. (PubMed)
88. Brown BG, Zhao XQ, Chait A, et al. Simvastatin and niacin, antioxidant vitamins, or the combination for the prevention of coronary disease. N Engl J Med. 2001;345(22):1583-1592. (PubMed)
89. Vittone F, Chait A, Morse JS, Fish B, Brown BG, Zhao XQ. Niacin plus simvastatin reduces coronary stenosis progression among patients with metabolic syndrome despite a modest increase in insulin resistance: a subgroup analysis of the HDL-Atherosclerosis Treatment Study (HATS). J Clin Lipidol. 2007;1(3):203-210. (PubMed)
90. Zhao XQ, Morse JS, Dowdy AA, et al. Safety and tolerability of simvastatin plus niacin in patients with coronary artery disease and low high-density lipoprotein cholesterol (The HDL Atherosclerosis Treatment Study). Am J Cardiol. 2004;93(3):307-312. (PubMed)
91. Sazonov V, Maccubbin D, Sisk CM, Canner PL. Effects of niacin on the incidence of new onset diabetes and cardiovascular events in patients with normoglycaemia and impaired fasting glucose. Int J Clin Pract. 2013;67(4):297-302. (PubMed)
92. Boden WE, Probstfield JL, Anderson T, et al. Niacin in patients with low HDL cholesterol levels receiving intensive statin therapy. N Engl J Med. 2011;365(24):2255-2267. (PubMed)
93. Michos ED, Sibley CT, Baer JT, Blaha MJ, Blumenthal RS. Niacin and statin combination therapy for atherosclerosis regression and prevention of cardiovascular disease events: reconciling the AIM-HIGH (Atherothrombosis Intervention in Metabolic Syndrome With Low HDL/High Triglycerides: Impact on Global Health Outcomes) trial with previous surrogate endpoint trials. J Am Coll Cardiol. 2012;59(23):2058-2064. (PubMed)
94. Kalil RS, Wang JH, de Boer IH, et al. Effect of extended-release niacin on cardiovascular events and kidney function in chronic kidney disease: a post hoc analysis of the AIM-HIGH trial. Kidney Int. 2015;87(6):1250-1257. (PubMed)
95. Landray MJ, Haynes R, Hopewell JC, et al. Effects of extended-release niacin with laropiprant in high-risk patients. N Engl J Med. 2014;371(3):203-212. (PubMed)
96. Schandelmaier S, Briel M, Saccilotto R, et al. Niacin for primary and secondary prevention of cardiovascular events. Cochrane Database Syst Rev. 2017;6:Cd009744. (PubMed)
97. Stone NJ, Robinson JG, Lichtenstein AH, et al. 2013 ACC/AHA guideline on the treatment of blood cholesterol to reduce atherosclerotic cardiovascular risk in adults: a report of the American College of Cardiology/American Heart Association Task Force on Practice Guidelines. J Am Coll Cardiol. 2014;63(25 Pt B):2889-2934. (PubMed)
98. Jackevicius CA, Tu JV, Ko DT, de Leon N, Krumholz HM. Use of niacin in the United States and Canada. JAMA Intern Med. 2013;173(14):1379-1381. (PubMed)
99. Burk K. Friedreich ataxia: current status and future prospects. Cerebellum Ataxias. 2017;4:4. (PubMed)
100. Chan PK, Torres R, Yandim C, et al. Heterochromatinization induced by GAA-repeat hyperexpansion in Friedreich's ataxia can be reduced upon HDAC inhibition by vitamin B3. Hum Mol Genet. 2013;22(13):2662-2675. (PubMed)
101. Libri V, Yandim C, Athanasopoulos S, et al. Epigenetic and neurological effects and safety of high-dose nicotinamide in patients with Friedreich's ataxia: an exploratory, open-label, dose-escalation study. Lancet. 2014;384(9942):504-513. (PubMed)
102. Lynch DR, Fischbeck KH. Nicotinamide in Friedreich's ataxia: useful or not? Lancet. 2014;384(9942):474-475. (PubMed)
103. Taylor EW. The oxidative stress-induced niacin sink (OSINS) model for HIV pathogenesis. Toxicology. 2010;278(1):124-130. (PubMed)
104. Favre D, Mold J, Hunt PW, et al. Tryptophan catabolism by indoleamine 2,3-dioxygenase 1 alters the balance of TH17 to regulatory T cells in HIV disease. Sci Transl Med. 2010;2(32):32ra36. (PubMed)
105. Jenabian MA, Patel M, Kema I, et al. Distinct tryptophan catabolism and Th17/Treg balance in HIV progressors and elite controllers. PLoS One. 2013;8(10):e78146. (PubMed)
106. Chen J, Shao J, Cai R, et al. Anti-retroviral therapy decreases but does not normalize indoleamine 2,3-dioxygenase activity in HIV-infected patients. PLoS One. 2014;9(7):e100446. (PubMed)
107. Dunham RM, Gordon SN, Vaccari M, et al. Preclinical evaluation of HIV eradication strategies in the simian immunodeficiency virus-infected rhesus macaque: a pilot study testing inhibition of indoleamine 2,3-dioxygenase. AIDS Res Hum Retroviruses. 2013;29(2):207-214. (PubMed)
108. Souza SA, Chow DC, Walsh EJ, Ford S, 3rd, Shikuma C. Pilot study on the safety and tolerability of extended release niacin for HIV-infected patients with hypertriglyceridemia. Hawaii Med J. 2010;69(5):122-125. (PubMed)
109. Dube MP, Lipshultz SE, Fichtenbaum CJ, et al. Effects of HIV infection and antiretroviral therapy on the heart and vasculature. Circulation. 2008;118(2):e36-40. (PubMed)
110. Carr A, Samaras K, Burton S, et al. A syndrome of peripheral lipodystrophy, hyperlipidaemia and insulin resistance in patients receiving HIV protease inhibitors. AIDS. 1998;12(7):F51-58. (PubMed)
111. Giannarelli C, Klein RS, Badimon JJ. Cardiovascular implications of HIV-induced dyslipidemia. Atherosclerosis. 2011;219(2):384-389. (PubMed)
112. Chow DC, Stein JH, Seto TB, et al. Short-term effects of extended-release niacin on endothelial function in HIV-infected patients on stable antiretroviral therapy. AIDS. 2010;24(7):1019-1023. (PubMed)
113. Balasubramanyam A, Coraza I, Smith EO, et al. Combination of niacin and fenofibrate with lifestyle changes improves dyslipidemia and hypoadiponectinemia in HIV patients on antiretroviral therapy: results of "heart positive," a randomized, controlled trial. J Clin Endocrinol Metab. 2011;96(7):2236-2247. (PubMed)
114. Dube MP, Komarow L, Fichtenbaum CJ, et al. Extended-release niacin versus fenofibrate in HIV-infected participants with low high-density lipoprotein cholesterol: effects on endothelial function, lipoproteins, and inflammation. Clin Infect Dis. 2015;61(5):840-849. (PubMed)
115. Hoffer LJ. Vitamin therapy in schizophrenia. Isr J Psychiatry Relat Sci. 2008;45(1):3-10. (PubMed)
116. Pauling L. Orthomolecular psychiatry. Varying the concentrations of substances normally present in the human body may control mental disease. Science. 1968;160(3825):265-271. (PubMed)
117. Seybolt SE. Is it time to reassess alpha lipoic acid and niacinamide therapy in schizophrenia? Med Hypotheses. 2010;75(6):572-575. (PubMed)
118. Zell M, Grundmann O. An orthomolecular approach to the prevention and treatment of psychiatric disorders. Adv Mind Body Med. 2012;26(2):14-28. (PubMed)
119. Yao JK, Dougherty GG, Jr., Gautier CH, et al. Prevalence and specificity of the abnormal niacin response: a potential endophenotype marker in schizophrenia. Schizophr Bull. 2016;42(2):369-376. (PubMed)
120. Sun L, Yang X, Jiang J, et al. Identification of the niacin-blunted subgroup of schizophrenia patients from mood disorders and healthy individuals in Chinese population. Schizophr Bull. 2017; doi: 10.1093/schbul/sbx150. [Epub ahead of print]. (PubMed)
121. Messamore E. Niacin subsensitivity is associated with functional impairment in schizophrenia. Schizophr Res. 2012;137(1-3):180-184. (PubMed)
122. Horrobin DF. The membrane phospholipid hypothesis as a biochemical basis for the neurodevelopmental concept of schizophrenia. Schizophr Res. 1998;30(3):193-208. (PubMed)
123. Messamore E. The niacin response biomarker as a schizophrenia endophenotype: A status update. Prostaglandins Leukot Essent Fatty Acids. 2017; pii: S0952-3278(16)30249-6. doi: 10.1016/j.plefa.2017.06.014. [Epub ahead of print]. (PubMed)
124. Jacob R, Swenseid M. Niacin. In: Ziegler E, Filer L, eds. Present Knowledge in Nutrition. 7th ed. Washington D.C.: ILSI Press; 1996:185-190.
125. US Department of Agriculture. USDA National Nutrient Database for Standard Reference, Release 25. 2012. https://ndb.nal.usda.gov/ndb/. Accessed 7/30/17.
126. Hendler SS, Rorvik DR. PDR for Nutritional Supplements. 2nd ed. Montvale: Thomson Reuters; 2008.
127. Minto C, Vecchio MG, Lamprecht M, Gregori D. Definition of a tolerable upper intake level of niacin: a systematic review and meta-analysis of the dose-dependent effects of nicotinamide and nicotinic acid supplementation. Nutr Rev. 2017;75(6):471-490. (PubMed)
128. MacKay D, Hathcock J, Guarneri E. Niacin: chemical forms, bioavailability, and health effects. Nutr Rev. 2012;70(6):357-366. (PubMed)
129. Dellinger RW, Santos SR, Morris M, et al. Repeat dose NRPT (nicotinamide riboside and pterostilbene) increases NAD(+) levels in humans safely and sustainably: a randomized, double-blind, placebo-controlled study. NPJ Aging Mech Dis. 2017;3:17. (PubMed)
130. Fulgoni VL, 3rd, Keast DR, Bailey RL, Dwyer J. Foods, fortificants, and supplements: Where do Americans get their nutrients? J Nutr. 2011;141(10):1847-1854. (PubMed)
131. Kar S, Chockalingam A. Statin-associated rhabdomyolysis with acute renal failure complicated by intradialytic NSTEMI: a review of lipid management considerations. Am J Ther. 2013;20(1):57-60. (PubMed)
132. Cziraky MJ, Willey VJ, McKenney JM, et al. Risk of hospitalized rhabdomyolysis associated with lipid-lowering drugs in a real-world clinical setting. J Clin Lipidol. 2013;7(2):102-108. (PubMed)
133. Maccubbin DL, Chen F, Anderson JW, et al. Effectiveness and safety of laropiprant on niacin-induced flushing. Am J Cardiol. 2012;110(6):817-822. (PubMed)
134. Hps Thrive Collaborative Group. HPS2-THRIVE randomized placebo-controlled trial in 25 673 high-risk patients of ER niacin/laropiprant: trial design, pre-specified muscle and liver outcomes, and reasons for stopping study treatment. Eur Heart J. 2013;34(17):1279-1291. (PubMed)
135. Lewey J, Shrank WH, Bowry AD, Kilabuk E, Brennan TA, Choudhry NK. Gender and racial disparities in adherence to statin therapy: a meta-analysis. Am Heart J. 2013;165(5):665-678, 678 e661. (PubMed)
136. Cheung MC, Zhao XQ, Chait A, Albers JJ, Brown BG. Antioxidant supplements block the response of HDL to simvastatin-niacin therapy in patients with coronary artery disease and low HDL. Arterioscler Thromb Vasc Biol. 2001;21(8):1320-1326. (PubMed)
137. Brown BG, Cheung MC, Lee AC, Zhao XQ, Chait A. Antioxidant vitamins and lipid therapy: end of a long romance? Arterioscler Thromb Vasc Biol. 2002;22(10):1535-1546. (PubMed)
138. Rios-Avila L, Coats B, Chi YY, et al. Metabolite profile analysis reveals association of vitamin B-6 with metabolites related to one-carbon metabolism and tryptophan catabolism but not with biomarkers of inflammation in oral contraceptive users and reveals the effects of oral contraceptives on these processes. J Nutr. 2015;145(1):87-95. (PubMed)
139. Dollerup OL, Christensen B,Svart M, et al. A randomized placebo-controlled clinical trial of nicotinamide riboside in obese men: safety, insulin-sensitivity, and lipid-mobilizing effects. Am J Clin Nutr. 2018;108:343-353. (PubMed)