Contenido
El cobre (Cu) es un elemento traza esencial para los seres humanos y animales. En el cuerpo, el cobre se desplaza entre sus formas cuproso (Cu1+) y cúprico (Cu2+), aunque la mayoría del cobre en el organismo se encuentra en forma (Cu2+). La habilidad del cobre para aceptar y donar electrones fácilmente explica la importancia de este mismo en las reacciones de oxidación y reducción (redox) y la actividad de barrido de radicales libres (1). Aunque se dice que Hipócrates prescribía compuestos de cobre para tratar enfermedades desde principios del 400 A.C. (2), científicos están aun descubriendo nueva información acerca de las funciones del cobre en el cuerpo humano (3).
El cobre es un componente crítico funcional de varias enzimas esenciales conocidas como cuproenzimas (4). Algunas de las funciones fisiológicas conocidas por ser cobre-dependientes son explicadas a continuación.
La enzima cobre-dependiente, llamada citocromo C oxidasa, juega un papel crítico en la producción de energía celular. Al catalizar la reducción del oxigeno molecular (O2) a agua (H2O), la citocromo c oxidasa genera un gradiente eléctrico usado por la mitocondria para crear la molécula de almacenaje energético vital, el ATP (5).
Otra cuproenzima, la lisil oxidasa, es necesaria para el entrecruzamiento del colágeno y la elastina, los cuales son esenciales para la formación de un tejido conjuntivo fuerte y flexible. La acción de la lisil oxidasa ayuda a mantener la integridad del tejido conjuntivo en el corazón y los vasos sanguíneos, y también participa en la formación ósea (2).
Cuatro enzimas que contienen cobre, conocidas como oxidasas multicobre o ferroxidasas tienen la capacidad de oxidar el hierro ferroso (Fe2+) en hierro férrico (Fe3+), la forma del hierro que puede ser cargada en la proteína transferrina para su transporte al sitio de formación de glóbulos rojos. La familia de oxidasas multicobre comprende la ceruloplasmina circulante (la cual representa ~90% del cobre en el plasma), la ceruloplasmina unida a la membrana (llamada ceruloplasmina GPI) y dos proteínas llamadas Hefaestina y Zyklopen encontradas en los intestinos y la placenta respectivamente (6). Interesantemente, ratones que no expresan ceruloplasmina (Cp-/-) tienen un metabolismo normal del cobre pero una acumulación anormal de hierro en el hígado (7, 8). Similarmente, individuos que necesitan ceruloplasmina muestran una sobrecarga de hierro en ciertos tejidos, incluyendo el hígado, cerebro, y la retina (9). Esto apoya la idea de que la actividad ferroxidasa de la ceruloplasmina es esencial para el flujo de hierro en el cuerpo. Por otra parte, el hecho de que la movilización del hierro de los lugares de almacenamiento se altera en la deficiencia de cobre apoya el papel de las oxidasas multicobre en el metabolismo del hierro (10).
Una serie de reacciones esenciales para el funcionamiento normal del cerebro y el sistema nervioso son catalizadas por cuproenzimas.
La dopamina β hidroxilasa cataliza la conversión de la dopamina para el neurotransmisor norepinefrina (11).
La envoltura de la mielina está hecha de fosfolípidos cuyas síntesis dependen de la actividad de la citocromo c oxidasa (2).
La cuproenzima, tirosinasa, es requerida para la formación del pigmento melanina. La melanina se forma en células llamadas melanocitos y juega un papel en la pigmentación del cabello, la piel y los ojos (2).
La enzima superóxido dismutasa (SOD) funciona como un antioxidante al catalizar la conversión de radicales superóxido (radicales libres) a peróxido de hidrógeno, el cual subsecuentemente puede ser reducido a agua por otras enzimas antioxidantes (12). Dos formas de la SOD contienen cobre: (1) la SOD con cobre/zinc se encuentra dentro de la mayoría de las células del cuerpo, incluyendo eritrocitos; y (2) la SOD extracelular que es una enzima contenedora de cobre encontrada en altos niveles en los pulmones y en bajos niveles de plasma sanguíneo (2).
La ceruloplasmina puede funcionar como un antioxidante de dos diferentes maneras. Iones libres de cobre y hierro son poderosos catalizadores del daño por radicales libres. Al unir el cobre, la ceruloplasmina previene que los iones libres de cobre catalicen el daño oxidativo. La actividad ferroxidasa de la ceruloplasmina (oxidación del hierro ferroso) facilita la carga del hierro en su proteína transportadora, la transferrina, y puede prevenir que los iones libres de hierro (Fe2+) participen en las reacciones dañinas que producen radicales libres (12).
Los niveles celulares del cobre pueden afectar la síntesis de las proteínas al potenciar o inhibir la transcripción de genes específicos. El cobre puede regular la expresión de los genes incrementando el nivel de estrés oxidativo intracelular. Un cierto número de vías de transducción de señales son activadas en respuesta al estrés oxidativo y pueden conducir al incremento en la expresión de genes envueltos en la desintoxicación de especies reactivas de oxigeno (13).
Un adecuado estatus nutricional del cobre es necesario para un metabolismo normal del hierro y para la formación de glóbulos rojos. La anemia es una señal clínica de la deficiencia del cobre, y el hierro ha sido encontrado acumulado en los hígados de animales con una deficiencia de cobre, indicando que el cobre (a través de la ceruloplasmina) es requerido para el transporte del hierro a la medula ósea para la formación de glóbulos rojos (ver Metabolismo del hierro) (2). La conexión entre la disponibilidad del cobre y el metabolismo del hierro ha sido también establecida en humanos; una deficiencia de cobre puede llevar a una deficiencia secundaria de ceruloplasmina y una sobrecarga de hierro hepático y/o cirrosis (10). La suplementación oral de cobre restauro los niveles de normales de ceruloplasmina y la actividad plasmática ferroxidasa y corrigió el desorden del metabolismo del hierro en un sujeto con deficiencia de cobre (14). Por otra parte, infantes alimentados con fórmulas altas en hierro absorbieron menos cobre que los infantes alimentados con fórmulas con bajos niveles de hierro, sugiriendo que altas ingestas de hierro en infantes pueden interferir con la absorción de cobre (15).
Altas ingestas suplementarias de zinc de 50 mg/día o más por periodos extensos de tiempo puede resultar en una deficiencia de cobre. Una alta ingesta dietaría de zinc incrementa la síntesis de una proteína celular llamada metalotioneína, la cual se une a ciertos metales y previene su absorción atrapándolos en las células intestinales. La metalotioneína tiene una afinidad superior al cobre que al zinc, es por ello que altos niveles of metalotioneína inducidos por un exceso de zinc causan un descenso en la absorción del cobre. En contraste, altas ingestas de cobre no se ha encontrado que afecten el estatus nutricional del zinc (2, 15). La suplementación con zinc (10 mg/día por 8 semanas) fue capaz de restaurar las proporciones normales de cobre/zinc del plasma en 65 sujetos con hemodiálisis a largo plazo que inicialmente exhibieron bajos niveles de zinc y elevados niveles de cobre. Si una mejora del estatus del zinc y el cobre en los pacientes de hemodiálisis puede impactar sus resultados clínicos aún necesita ser evaluada (16).
Dietas altas en fructosa han exacerbado la deficiencia de cobre en ratas pero no en cerdos los cuales poseen un sistema gastrointestinal más parecido al de los seres humanos. Muy altos niveles de fructuosa dietaría (20% del total de las calorías) no resultaron en una reducción de cobre en humanos, sugiriendo que la ingesta de fructosa no resulta en una reducción del cobre en niveles relevantes de dietas normales (2, 15).
Aunque los suplementos de vitamina C han producido deficiencia de cobre en conejillos de indias (17), el efecto de los suplementos de vitamina C en el estado nutricional del cobre en seres humanos es menos claro. Dos estudios menores en hombres adultos jóvenes sanos indican que la actividad oxidasa de la ceruloplasmina puede verse deteriorada por dosis relativamente altas de vitamina C suplementaria. En un estudio, la suplementación con vitamina C de 1,500 mg/día por dos meses resultó en una disminución significativa de la actividad oxidasa de la ceruloplasmina (18). En el otro estudio, los suplementos de 605 mg de vitamina C/día por tres semanas provocaron una disminución en la actividad oxidasa de la ceruloplasmina, aunque la absorción de cobre no disminuyo (19). Ninguno de estos estudios encontró que la suplementación de vitamina C afectara adversamente el estatus nutricional del cobre.
La deficiencia franca del cobre dietario o clínicamente evidente es relativamente poco común. Los niveles del cobre en el suero y la ceruloplasmina pueden caer hasta un 30% de lo normal en casos de severa deficiencia del cobre. La hipocupremia (bajo contenido de cobre en la sangre) es también observada en desordenes genéticos debido al metabolismo del cobre como la aceruloplasminemia y paradójicamente en la enfermedad de Wilson, los cuales no están ligados a una deficiencia dietaría del cobre. Uno de los signos clínicos más comunes de la deficiencia del cobre es una anemia que no responde a la terapia con hierro pero es corregida con una suplementación de cobre. Aunque se piensa que la anemia es el resultado defectuoso de la movilización del hierro debido a la disminución de la actividad de la ceruloplasmina, la ausencia de ceruloplasmina en individuos con aceruloplasminemia hereditaria no está siempre asociada con la anemia manifiesta (20). La deficiencia del cobre puede conducir también a anormales bajos números de glóbulos blancos conocidos con neutrófilos (neutropenia), una condición que puede ser acompañada por un incremento en la susceptibilidad a infecciones. Estudios de repleción de cobre han sugerido que la reducción en la disponibilidad del cobre puede afectar el linaje de las células eritroides y mieloides, apoyando el papel del cobre en la regulación de la renovación de células sanguíneas (21, 22). Más investigación es claramente necesaria para descubrir los mecanismos subyacentes de la deficiencia inducida del cobre en la anemia y neutropenia (4, 23). La osteoporosis y otras anormalidades del desarrollo óseo relacionados con la deficiencia del cobre son los más comunes en lactantes de bajo peso al nacer con deficiencia de cobre y niños pequeños. Características menos comunes de la deficiencia de cobre puede incluir perdida de la pigmentación, síntomas neurológicos, y problemas de crecimiento (2, 5).
La leche de vaca es relativamente baja en cobre, y se han reportado casos de deficiencia de cobre de alto riesgo en infantes y niños alimentados sólo con fórmula de leche de vaca (24). Individuos de alto riesgo incluyen infantes prematuros (especialmente aquellos con bajo peso al nacer), infantes con diarrea prolongada, infantes y niños en recuperación de malnutrición, e individuos con síndromes de malabsorción, incluyendo enfermedad celiaca, esprúe, y síndrome del intestino corto debido a una remoción quirúrgica de una gran porción del intestino. Los individuos que reciben nutrición parenteral total intravenosa carente de cobre u otras dietas restringidas, pueden también necesitar de la suplementación con cobre y otros elementos trazas (2, 5). La deficiencia de cobre en infantes con colestasis (reducida excreción biliar del cobre) ha sido ligada a la nutrición parental a largo plazo carente de cobre (25). Reportes de casos indican que los pacientes con fibrosis quística pueden también estar en mayor riesgo de una deficiencia de cobre (26). Finalmente, una ingesta excesiva de zinc ha llevado a una deficiencia de cobre secundaria en individuos al usar suplementos de zinc o cremas dentales enriquecidas con zinc (27, 28).
Un síndrome neurológico ha sido descrito en adultos con una deficiencia de cobre adquirida (29). Los síntomas incluyen desmielinización, mielopatía, y polineuropatia del sistema nervioso central y una inflamación del nervio óptico. La etiología es desconocida en ausencia de factores de riesgo prominentes (vea factores de riesgo en la siguiente sección: Individuos en riesgo de deficiencia); reportes de casos describen un incremento del contenido de cobre en los intestinos sugiriendo un síndrome de mal absorción como la enfermedad de Menkes, pero las mutaciones en el gen ATP7 no estuvieron ligadas a la condición (vea Deficiencia de cobre hereditaria en la parte de abajo) (30). El remplazo oral de cobre (2 mg/día de cobre elemental) normaliza las concentraciones de cobre y ceruloplasmina en el suero, estabiliza la condición y mejora significativamente la calidad de vida de los sujetos afectados. Sin embargo, la duración de la suplementación con cobre no ha sido establecida aun, y la incrementación de la dosis puede ser requerida en caso de una recaída (29).
El trafico del cobre dentro de la mayoría de la células excepto por los hepatocitos (células del hígado) esta facilitado por el transporte de Cu1+- y ATPasa llamada ATP7A. Mutaciones en el gen ATP7A perjudican el transporte de cobre intracelular, el cual se acumula en el citosol de los enterocitos y las células endoteliales vasculares (31). Esto resulta en una deficiencia sistémica de cobre y disminuye la actividad de las cuproenzimas. El transporte del cobre en el cerebro es también afectado, llevando a una acumulación de cobre en la barrera hematoencefálica del cerebro y a una disminución de la actividad de las cuproenzimas en las neuronas. Los individuos afectados son diagnosticados con la enfermedad de Menkes o con una forma más leve de la enfermedad llamada síndrome del cuerno occipital (OHS por sus siglas en Inglés). Las características clínicas de la enfermedad de Menkes (MD) incluyen convulsiones intratables, desordenes del tejido conjuntivo, hemorragia subdural, y anormalidades del cabello ("cabello ensortijado"). Los pacientes con OHS exhiben hipotonía muscular y anormalidades del tejido conjuntivo, incluyendo exostosis en los huesos occipitales. Inyecciones subcutáneas de cobre-histidina son usadas para evitar la absorción intestinal defectuosa y mejorar las funciones metabólicas del cobre en los pacientes. Sin embargo, la entrada de cobre en el cerebro sigue siendo limitada (revisado en 32).
Una variedad de indicadores fue usada para establecer la IDR para el cobre, incluyendo la concentración de cobre en el plasma, la actividad de ceruloplasmina plasmática, la actividad de la superóxido dismutasa en los eritrocitos, y las concentraciones de cobre plaquetario (15). La IDR de cobre refleja los resultados de estudios de eliminación y reposición, y está basada en la prevención de una deficiencia.
| Etapa de la Vida | Edad | Machos (μg/día) | Hembras (μg/día) |
|---|---|---|---|
| Infantes | 0-6 meses | 200 (IA) | 200 (IA) |
| Infantes | 7-12 meses | 220 (IA) | 220 (IA) |
| Niños | 1-3 años | 340 | 340 |
| Niños | 4-8 años | 440 | 440 |
| Niños | 9-13 años | 700 | 700 |
| Adolescentes | 14-18 años | 890 | 890 |
| Adultos | 19 años y más | 900 | 900 |
| Embarazo | Todas las edades | - | 1,000 |
| Período de lactancia | Todas las edades | - | 1,300 |
Mientras está claro que la severa deficiencia de cobre resulta en anormalidades y daño cardíaco (cardiomiopatía) en algunas especies de animales, la patología difiere de la enfermedad cardiovascular aterosclerótica que prevalece en seres humanos (15). Estudios en humanos han producido resultados inconsistentes, y su interpretación se dificulta por la falta de un marcador confiable del estado nutricional del cobre. Fuera del cuerpo, el cobre que se encuentra libre es conocido por ser un pro-oxidante y es frecuentemente usado para producir la oxidación de lipoproteínas de baja densidad (LDL) en tubos de ensayos. Se ha encontrado que la proteína contenedora de cobre, la ceruloplasmina estimula la oxidación de proteínas de baja densidad en tubos de ensayo (33), llevando a algunos científicos a proponer que el incremento en los niveles de cobre podría incrementar el riesgo de aterosclerosis al promover la oxidación de LDL. Sin embargo, hay poca evidencia de que el cobre o la ceruloplasmina promuevan la oxidación de LDL en el cuerpo humano. Además, las cuproenzimas superóxido dismutasa y ceruloplasmina, son conocidas por tener propiedades antioxidantes, llevando a que algunos expertos propongan que más bien es la deficiencia de cobre en vez del exceso la que incrementa el riesgo de enfermedades cardiovasculares (34).
Varios estudios epidemiológicos han encontrado que el incremento en los niveles de cobre en el plasma se asocia con un incremento del riesgo de padecer de enfermedades cardiovasculares. Un estudio de cohorte prospectivo en los EE. UU. examinó los niveles de cobre en el plasma en más de 4,500 hombres y mujeres de entre 30 o más años de edad (35). Durante 16 años de seguimiento, 151 participantes murieron de enfermedad coronaria. Después del ajuste por otros factores de riesgo de enfermedades cardíacas, aquellos con niveles de cobre plasmático en los dos cuartiles más altos tuvieron un riesgo significativamente mayor de morir de una enfermedad coronaria. Otros tres estudios de caso y control llevados a cabo en Europa tuvieron hallazgos similares. Un estudio menor en 60 pacientes con falla cardíaca crónica o enfermedad cardíaca isquémica reportó que el cobre del suero era un predictor de resultados a corto plazo (36). Otro estudio de cohorte prospectivo en 4,035 hombres de mediana edad reportó que altos niveles de cobre en el suero estaban significativamente relacionados con un incremento del 50% en la mortalidad general; sin embargo, el cobre plasmático no estuvo asociado significativamente con la mortalidad cardiovascular en este estudio (37). Adicionalmente, se ha encontrado que el cobre en el suero es elevado en pacientes con cardiopatía reumática (38).
Es importante notar que el cobre en el suero refleja en gran medida la ceruloplasmina y no es un indicador sensible del estado nutricional del cobre (39). Los niveles de ceruloplasmina plasmática son conocidos por incrementar en un 50% o más, bajo ciertas condiciones de estrés físico, como traumas, inflamación o enfermedades. Debido a que más del 90% del cobre plasmático es llevado en la ceruloplasmina, la cual se incrementa en condiciones inflamatorias, el cobre plasmático elevado puede ser simplemente un marcador de la inflamación que acompaña la aterosclerosis. En contraste a los hallazgos epidemiológicos que vinculan el incremento de los niveles de cobre en el suero con la enfermedad cardíaca, dos estudios de autopsias encontraron que los niveles de cobre en el músculo cardíaco eran en realidad más bajos en pacientes que murieron de enfermedad coronaria que en aquellos que murieron por otras causas (40). Además, el contenido de cobre en los glóbulos blancos ha sido positivamente correlacionado con el grado de permeabilidad de las arterias coronarias en pacientes con enfermedad coronaria (41, 42). Aún más, los pacientes con historia de infarto al miocardio (IM) tuvieron concentraciones de superóxido dismutasa extracelular más bajas que aquellos sin un historial de IM (43). Así, debido a la carencia de un biomarcador confiable del estado nutricional del cobre, no es claro si el cobre está o no relacionado con las enfermedades cardiovasculares.
Mientras que algunos estudios en un número muy reducido de adultos, alimentados con dietas experimentales bajas en cobre, han demostrado cambios adversos en los niveles de colesterol sanguíneo, incluyendo un incremento total en los niveles de colesterol LDL, y una disminución en los niveles de colesterol HDL (44), otros estudios no han confirmado estos resultados (45). La suplementación de cobre de 2-3 mg/día por cuatro u ocho semanas no resulto en cambios clínicos significantes en los niveles de colesterol (34, 46, 47). Además, la investigación ha fallado en encontrar evidencia que el incremento en la ingesta de cobre incremente el estrés oxidativo. En una prueba multicéntrica controlada con placebo, la suplementación con cobre de 3 y 6 mg/día por seis semanas no resultó en un incremento de la susceptibilidad a la oxidación de LDL, inducida fuera del cuerpo (ex vivo) por cobre o peroxinitrito (una especie reactiva del nitrógeno) (48). Más aún, la suplementación con 3 y 6 mg/día de cobre disminuyó la oxidabilidad in vitro de glóbulos rojos (49), indicando que las ingestas de cobre relativamente altas no incrementan la susceptibilidad a de LDL o de glóbulos rojos a la oxidación.
Aunque el cobre libre y la ceruloplasmina pueden promover la oxidación de LDL en tubos de ensayo, hay poca evidencia de que un incremento de cobre dietario aumente el estrés oxidativo en el cuerpo humano. El incremento en los niveles de cobre en el suero han sido asociados con un incremento del riesgo de enfermedades cardiovasculares, pero la importancia de estos hallazgos no está clara debido a la asociación entre los niveles de ceruloplasmina en la sangre y las condiciones inflamatorias. La aclaración de las relaciones entre el estado nutricional del cobre, los niveles de ceruloplasmina y el riesgo de enfermedades cardiovasculares, requiere de más investigación.
Notablemente, se sugirió que las concentraciones elevadas de cobre en el plasma pudieran estar ligadas a niveles altos de homocisteína en individuos con enfermedades vasculares (50, 51). Los niveles incrementados de homocisteína están asociados con lesiones de la pared arterial y un incremento en el riesgo de padecer enfermedades cardiovasculares (52). La interacción entre la homocisteína y el cobre fue ligada a una alteración en la función endotelial vascular en modelos animales (53, 54). Sin embargo, aunque la restricción de cobre en animales ha mostrados algunos efectos benéficos en los niveles de homocisteína y lesiones aterogénicas (55, 56), aun no se conoce si el desequilibrio del cobre contribuye al efecto aterogénico de la homocisteína en seres humanos.
Se sabe que el cobre juega un papel importante en el desarrollo y mantenimiento de la función del sistema inmune, pero el mecanismo exacto de su acción aún se desconoce. La neutropenia (cantidad anormalmente baja de los glóbulos blancos conocidos como neutrófilos) es un signo clínico de la deficiencia de cobre en seres humanos. Los efectos adversos por insuficiencia de cobre en el sistema inmune parecen ser más pronunciados en infantes. Infantes con síndrome de Menkes, un desorden genético que resulta en una deficiencia de cobre severa, sufren de frecuentes infecciones severas (57, 58). En un estudio de 11 niños malnutridos con evidente deficiencia de cobre, la habilidad de ciertos glóbulos blancos de fagocitar patógenos incrementó significativamente después de un mes de suplementación con cobre (59). Por otra parte, 11 hombres con una dieta baja en cobre (0.66 mg de cobre/día por 24 días y 0.38 mg de cobre/día por otros 40 días) mostraron una disminución de la respuesta proliferaría cuando glóbulos blancos denominados monocitos fueron aislados de su sangre, y presentados con un reto inmunológico en cultivo celular (60). Recientes estudios mecanicistas apoyan el papel del cobre en la innata respuesta inmune contra infecciones bacterianas (examinado en 61). Mientras que una severa deficiencia de cobre tiene efectos adversos en la función inmune, los efectos de la insuficiencia de cobre marginal en seres humanos no están todavía claros.
La pérdida progresiva de la densidad mineral ósea (DMO) llevando a una osteopenia (pre-osteoporosis) y osteoporosis es comúnmente observada en la población de adultos mayores. Las mujeres son más afectadas por la osteoporosis que los hombres (ej. una razón de 5:1 en blancos no hispánicos) (62), principalmente debido a la reducción de la producción de estrógeno en la postmenopausia que es esencial para el mantener la fuerza en músculos, huesos, y tejido conectivo (63). La osteoporosis está asociada con un incremento del riesgo de caídas, fracturas de huesos, y mortalidad en individuos mayores de 65 años de edad (64). La osteoporosis también ha sido reportada en infantes con una severa deficiencia de cobre (65, 66) pero no está completamente claro si la deficiencia de cobre marginal durante la adultez contribuye a desarrollar osteoporosis. Mientras que un incremento en la resorción ósea fue observada en 11 adultos masculinos sanos con una ingesta de cobre marginal de 0.7 mg/día por 6 semanas (67), la suplementación de 3 a 6 mg/día de cobre por seis semanas no tuvo efecto en los marcadores bioquímicos de la resorción ósea o formación ósea en dos estudios en hombres y mujeres adultos sanos (68, 69). Sin embargo, es posible que una reducción de la ingesta de cobre y absorción en adultos mayores reduzca la actividad de la enzima cobre-dependiente lisil oxidasa, la cual es requerida en la maduración del colágeno — un elemento clave en la matriz orgánica del hueso.
En general, la investigación con respecto al papel del estado nutricional del cobre en la osteoporosis senil es limitada. Un estudio temprano encontró que los niveles de cobre en el suero en 46 pacientes mayores con fracturas en la cadera fueron significativamente más bajos que aquellos de control emparejado (70). Sin embargo, otro estudio no encontró diferencias en los niveles de cobre en el suero entre mujeres postmenopáusicas con una DMO normal (N=40), osteopenia (N=40) u osteoporosis (N=40) (71). Un pequeño estudio en mujeres perimenopáusicas, que consumieron diariamente un promedio de 1 mg de cobre dietario, reportó una disminución en la pérdida de densidad mineral ósea (DMO) de la columna lumbar luego de la suplementación con cobre de 3 mg/día por dos años (72). Adicionalmente, un ensayo doble ciego controlado con placebo de dos años, en 59 mujeres postmenopáusicas, encontró que la combinación de calcio suplementario y minerales trazas, incluyendo 2.5 mg diarios de cobre, resultó en la mantención de la densidad ósea de la columna dorsal, mientras que el calcio suplementario o los minerales trazas por sí solos, no fueron efectivos en la prevención de la pérdida de la densidad ósea (73). Sin embargo, un estudio aleatorio, doble ciego controlado con placebo más reciente el cual inicialmente se llevó a cabo en 224 mujeres postmenopáusicas sanas de entre 51 a 80 años de edad encontró que la suplementación diaria de 600 mg de calcio, 2 mg de cobre, y 12 mg de zinc por 2 años disminuyo la densidad mineral ósea (DMO) de todo el cuerpo comparada con la suplementación de calcio solo. Más aun, aunque la DMO fue claramente reducida en sujetos con ingestas dietarías de cobre debajo de la IDR (0.9 mg/día), el cobre suplementario no previno la pérdida progresiva de la DMO así como el régimen de calcio solo (74). Finalmente varios estudios han sugerido que la perdida de dientes puede estar relacionada a un DMO sistémico pobre (75, 76). Cuando se comparó con 20 controles sanos emparejados, 50 pacientes (con un promedio de 47.5 años) con una DMO baja en la espina dorsal y un desgate dental avanzado fueron encontrados con un significativo contenido bajo de cobre en el esmalte de los dientes. Sin embargo, a pesar de que evidencia sugiere una desmineralización ósea, los niveles de cobre en el suero en esta población fueron similares a aquellos del grupo sano (77). Más investigación es requerida para llegar a una conclusión con respecto a los efectos de la deficiencia de cobre marginal, y la suplementación de cobre en el metabolismo óseo y la osteoporosis senil.
El deterioro cognitivo en individuos con la enfermedad de Alzheimer está ligado a la presencia de placas de β-amiloide y agregados anormales de proteínas formadoras de Tau. La posibilidad de que el desequilibrio del cobre está envuelto en el comienzo del Alzheimer está actualmente siendo investigado. En primer lugar parece que la fracción de cobre libre (no unido a la ceruloplasmina) esta aumentada en trastornos de la homeostasis de cobre, como también en los individuos con Alzheimer (78, 79). Más aun, pacientes con Alzheimer parecen tener altos niveles de cobre plasmático comparados con los que tiene un control sano (80). Entre las muchas hipótesis que apoyan el papel del cobre en el comienzo o progresión del Alzheimer, se sugirió que el cobre podía estar involucrado en la formación de placas seniles a través de la hipermetilación de los péptidos β-amiloides posiblemente llevando a una depleción del zinc, un mayor estrés oxidativo e incluso daño cerebral (81, 82). Investigaciones recientes han identificado también variaciones genéticas (polimorfismo) en el gen ATP7B que puede modificar el riesgo de desarrollar Alzheimer (83). La proteína, ATP7B, es responsable por la excreción de cobre hepático en el tracto biliar, y su deterioro en la enfermedad de Wilson resulta en un incremento en el nivel de cobre “libre” en la sangre y la acumulación de cobre en el cerebro y el hígado.
Más investigaciones son necesarias para indagar si las variantes genéticas pudieran influenciar la susceptibilidad de una exposición ambiental a altos niveles de cobre. La adición de cobre al agua potable ha sido asociada con características patológicas mejoradas en modelos animales con Alzheimer (84, 85). Un estudio en un conejo reporto que combinando una alta dieta de alto colesterol y cobre (0.12 mg/L en agua bebible) pudo poner en peligro la cognición (84). Un estudio de cohorte prospectivo en 3,718 participantes de edad avanzada del Chicago Health and Aging Project, con un seguimiento de 5.5 años, evaluó el impacto de las ingestas de ácidos grasos y cobre usando cuestionarios de frecuencia alimentaria y varias evaluaciones cognitivas. Para los individuos un consumo alto de ácidos grasos saturados y ácidos grasos trans el deterioro cognitivo fue mayor para aquellos en el quintil más elevado del total del consumo de cobre comparado con aquellos en el quintil más bajo (ingesta promedio de 2.75 vs 0.88 mg/día) (86).
Aunque un metabolismo de cobre disfuncional es sugerido como un factor de riesgo para el Alzheimer, podría ser también un síntoma de la enfermedad. Por otra parte, no es claro aun si la suplementación o restricción del cobre podría retrasar la progresión del Alzheimer. Una pequeña prueba contralada doble ciega con placebo en 68 individuos con Alzheimer en un estado leve encontró que la suplementación de 8 mg/día de cobre por un año retrasó la disminución del péptido β-amiloide Aβ42 en el fluido cerebroespinal; una disminución en Aβ42 ha sido ligado a un deterioro cognitivo (87). Sin embargo, este retraso no estuvo asociado con una mejora en el rendimiento cognitivo (88). Basado en el utilización de agentes quelantes de cobre en la enfermedad de Wilson, el reciente uso de acetato de zinc de liberación lenta (150 mg/día por 6 meses) en un estudio aleatorio controlado con placebo en 60 pacientes con leve y Alzheimer moderado resulto en una disminución de cobre libre en el suero y la estabilización de los déficits de la cognición (81). Estudios adicionales en humanos son necesarios para clarificar el papel del cobre en el comienzo y progresión del Alzheimer, y evaluar si el cobre dietario puede ayudar a prevenir Alzheimer en individuos con alto riesgo o controlar la enfermedad en pacientes con Alzheimer.
Tanto la aparición neurológica de la enfermedad de Wilson y la aceruloplasminemia hereditaria ambas están caracterizadas por la acumulación de cobre en el cerebro, resultando en síntomas neurológicos (distonía y deterioro cognitivo) que se parecen a la enfermedad de Parkinson (89). El nivel de cobre es disminuido en regiones cerebrales de perdida neuronal en pacientes con Parkinson (90). Sim embargo, el reciente metaánalisis de estudios que midieron los niveles de cobre en el suero, plasma y liquido cefalorraquídeo no encontraron ninguna diferencia entre pacientes con Parkinson y sujetos de edad avanzada sanos (91).
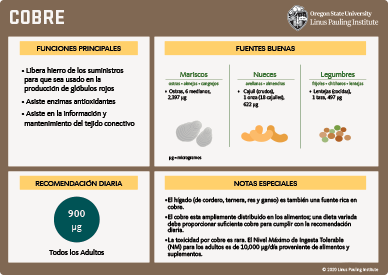
El cobre se encuentra en una amplia variedad de alimentos y es más abundante en vísceras, mariscos, frutos secos, y semillas. Los cereales de salvado de trigo y los productos de grano entero son también buenas fuentes de cobre. De acuerdo a encuestas nacionales, la ingesta dietética de cobre promedio en los EE.UU. es aproximadamente de 1.0 a 1.1 mg (1,000 a 1,100 μg)/día para mujeres adultas y de 1.2 a 1.6 mg (1,200 a 1,600 μg)/día para hombres adultos (15). El contenido de cobre de algunos alimentos que son relativamente ricos en cobre se muestra en microgramos (μg) en la tabla a continuación. Para mayor información sobre el contenido de nutrientes de los alimentos, revise base de datos de composición de los alimentos de la USDA.
| Alimento | Porción | Cobre (μg) |
|---|---|---|
| Hígado (res), cocido, frito | 1 onza | 4,128 |
| Moluscos, ostras, cocidas | 6 ostras medianas | 2,397 |
| Carne de cangrejo, cocida | 3 onzas | 1,005 |
| Carne de cangrejo, azul, cocida, asada | 3 onzas | 692 |
| Moluscos, almejas, especies mixtas, cocidas, asadas | 3 onzas | 585 |
| Castaña de cajú | 1 onza | 622 |
| Semillas de girasol, secas, tostadas | 1 onza | 519 |
| Avellanas, secas tostadas | 1 onza | 496 |
| Almendras | 1 onza | 292 |
| Mantequilla de maní, sin sal | 2 cucharadas | 185 |
| Lentejas, semillas maduras, cocidas, hervidas, sin sal | 1 taza | 497 |
| Champiñones, blancos, crudos | 1 taza (rebanados) | 223 |
| Cereal de trigo molido | 2 galletas | 167 |
| Chocolate (semidulce) | 1 onza | 198 |
Suplementos de cobre están disponibles como oxido cúprico, gluconato de cobre, sulfato de cobre, quelatos aminoácidos de cobre (92).
La toxicidad del cobre es poco común en la población general. El envenenamiento agudo con cobre ha ocurrido a través de la contaminación de bebidas que son almacenadas en contenedores con contenido de cobre, así como de suministros de agua contaminada (93). En los EE.UU. la pauta sanitaria para la máxima concentración de cobre en el agua potable es impuesta por la Agencia de Protección Medioambiental Estadounidense (1.3 mg/litro) y por la Organización Mundial de la Salud (OMS) (2 mg/litro) (94). Los síntomas de toxicidad aguda por cobre incluyen dolor abdominal, nauseas, vómitos y diarrea, los cuales ayudan a prevenir una ingesta y absorción adicional de cobre. Signos más graves de toxicidad aguda por cobre incluyen daño hepático severo, falla renal, coma, y muerte. Más preocupante desde un punto de vista nutricional, es la posibilidad de un daño hepático como resultado de una exposición a largo plazo a dosis de cobre más bajas. En individuos generalmente sanos, las dosis de hasta 10,000 μg (10 mg) diarios no han resultado en un daño hepático. Por esta razón, la Junta de Nutrición y Alimentos de los EE.UU. estableció el nivel máximo de ingesta tolerable (NM) de cobre de alimentos y suplementos en 10 mg/día (15). Se debe destacar que los individuos con desórdenes genéticos que afectan al metabolismo del cobre (ej., enfermedad de Wilson, cirrosis infantil de la India, y toxicosis por cobre idiopática) pueden estar en riesgo de efectos adversos de la toxicidad crónica por cobre a niveles de ingesta significativamente más bajos. Existe un poco de preocupación de que la NM de 10 mg/día pueda ser muy alta. En particular, en un estudio hombres consumieron 7.8 mg/día de cobre por 147 días. Ellos acumularon cobre durante ese lapso de tiempo, y algunos índices de la función inmune y antioxidante sugieren que esas funciones fueron adversamente afectadas por las altas ingestas de cobre (95, 96). Sin embargo, otro estudio no reporto algún efecto adverso en individuos suplementados con 8 mg/día de cobre por 6 meses (88).
| Grupo Etario | NM (μg/día) |
|---|---|
| Infantes 0-12 meses | Imposible de establecer* |
| Niños 1-3 años | 1,000 |
| Niños 4-8 años | 3,000 |
| Niños 9-13 años | 5,000 |
| Adolescentes 14-18 años | 8,000 |
| Adultos 19 años y más | 10,000 |
| *La fuente de la ingesta debiera ser sólo de alimentos y fórmula. | |
Relativamente poco es lo que se conoce acerca de la interacción del cobre con drogas. La penicilamina se utiliza para unir cobre y aumentar su eliminación en la enfermedad de Wilson, un desorden genético que provoca una sobrecarga de cobre hepático. Debido a que la penicilamina incrementa dramáticamente la excreción urinaria de cobre, los individuos que toman este medicamento por razones distintas a la sobrecarga de cobre, pueden tener un incremento del requerimiento de cobre. Adicionalmente, los antiácidos pueden intervenir con la absorción del cobre cuando se utilizan en muy altas cantidades (2).
La IDR del cobre (900 μg/día en adultos) es suficiente para prevenir una deficiencia, pero la falta de indicadores claros del estado nutricional del cobre en seres humanos, hace difícil determinar el nivel de ingesta de cobre más adecuado para promover una salud óptima o prevenir enfermedades crónicas. Una dieta variada debería aportar suficiente cobre a la mayoría de las personas. Para aquellos preocupados de que su dieta pueda no aportar el cobre suficiente, un suplemento multivitamínico/mineral generalmente aportará, al menos, la IDR de cobre.
Debido a que el envejecimiento no se ha asociado con cambios significativos en los requerimientos de cobre, nuestra recomendación para los adultos mayores es la misma que para los adultos de 50 años o menos (97).
Escrito en Abril de 2003 por:
Jane Higdon, Ph.D.
Instituto Linus Pauling
Universidad Estatal de Oregon
Actualizado en Julio de 2007 by:
Victoria J. Drake, Ph.D.
Instituto Linus Pauling
Universidad Estatal de Oregon
Actualizado en Diciembre de 2013 por:
Barbara Delage, Ph.D.
Instituto Linus Pauling
Universidad Estatal de Oregon
Revisado en Enero de 2014 por:
Joseph R. Prohaska, Ph.D.
Profesor de Bioquímica, Emérito
Universidad de Minnesota Escuela de Medicina Duluth
Traducido al Español en 2015 por:
Silvia Vazquez Lima
Instituto Linus Pauling
Universidad Estatal de Oregon
Originalmente traducido al español en 2012 por Guillermo Sandoval y editado por Andrew Quest (Ph.D.) y Lisette Leyton (Ph.D.), todos provenientes de la Universidad de Chile. Estos esfuerzos fueron patrocinados por el projecto Anillo #ACT1111, CONICYT-Chile, programa PIA.
Derechos de autoría 2001-2024 Instituto Linus Pauling
1. Linder MC, Hazegh-Azam M. Copper biochemistry and molecular biology. Am J Clin Nutr. 1996;63(5):797S-811S. (PubMed)
2. Turnlund JR. Copper. In: Shils ME, Shike M, Ross AC, Caballero B, Cousins RJ, eds. Modern Nutrition in Health and Disease. 10th ed. Philadelphia: Lippincott Williams & Wilkins; 2006:286-299.
3. Prohaska JR. Copper. In: Erdman JW, Macdonald IA, Zeisel SH, eds. Present Knowledge in Nutrition. 10th ed. Ames: Wiley-Blackwell; 2012:540-553.
4. Prohaska JR. Impact of copper limitation on expression and function of multicopper oxidases (ferroxidases). Adv Nutr. 2011;2(2):89-95. (PubMed)
5. Uauy R, Olivares M, Gonzalez M. Essentiality of copper in humans. Am J Clin Nutr. 1998;67(5 Suppl):952S-959S. (PubMed)
6. Vashchenko G, MacGillivray RT. Multi-copper oxidases and human iron metabolism. Nutrients. 2013;5(7):2289-2313. (PubMed)
7. Meyer LA, Durley AP, Prohaska JR, Harris ZL. Copper transport and metabolism are normal in aceruloplasminemic mice. J Biol Chem. 2001;276(39):36857-36861. (PubMed)
8. Harris ZL, Durley AP, Man TK, Gitlin JD. Targeted gene disruption reveals an essential role for ceruloplasmin in cellular iron efflux. Proc Natl Acad Sci U S A. 1999;96(19):10812-10817. (PubMed)
9. Kono S. Aceruloplasminemia. Curr Drug Targets. 2012;13(9):1190-1199. (PubMed)
10. Thackeray EW, Sanderson SO, Fox JC, Kumar N. Hepatic iron overload or cirrhosis may occur in acquired copper deficiency and is likely mediated by hypoceruloplasminemia. J Clin Gastroenterol. 2011;45(2):153-158. (PubMed)
11. Harris ED. Copper. In: O'Dell BL, Sunde RA, eds. Handbook of nutritionally essential minerals. New York: Marcel Dekker, Inc; 1997:231-273.
12. Johnson MA, Fischer JG, Kays SE. Is copper an antioxidant nutrient? Crit Rev Food Sci Nutr. 1992;32(1):1-31.
13. Mattie MD, McElwee MK, Freedman JH. Mechanism of copper-activated transcription: activation of AP-1, and the JNK/SAPK and p38 signal transduction pathways. J Mol Biol. 2008;383(5):1008-1018. (PubMed)
14. Videt-Gibou D, Belliard S, Bardou-Jacquet E, et al. Iron excess treatable by copper supplementation in acquired aceruloplasminemia: a new form of secondary human iron overload? Blood. 2009;114(11):2360-2361. (PubMed)
15. Food and Nutrition Board, Institute of Medicine. Copper. Dietary referencia intakes for vitamin A, vitamin K, boron, chromium, copper, iodine, iron, manganese, molybdenum, nickel, silicon, vanadium, and zinc. Washington, D.C.: National Academy Press; 2001:224-257. (National Academy Press)
16. Guo CH, Wang CL. Effects of zinc supplementation on plasma copper/zinc ratios, oxidative stress, and immunological status in hemodialysis patients. Int J Med Sci. 2013;10(1):79-89. (PubMed)
17. Milne DB, Omaye ST. Effect of vitamin C on copper and iron metabolism in the guinea pig. Int J Vitam Nutr Res. 1980;50(3):301-308. (PubMed)
18. Finley EB, Cerklewski FL. Influence of ascorbic acid supplementation on copper status in young adult men. Am J Clin Nutr. 1983;37(4):553-556. (PubMed)
19. Jacob RA, Skala JH, Omaye ST, Turnlund JR. Effect of varying ascorbic acid intakes on copper absorption and ceruloplasmin levels of young men. J Nutr. 1987;117(12):2109-2115. (PubMed)
20. Harris ZL, Klomp LW, Gitlin JD. Aceruloplasminemia: an inherited neurodegenerative disease with impairment of iron homeostasis. Am J Clin Nutr. 1998;67(5 Suppl):972S-977S. (PubMed)
21. Bustos RI, Jensen EL, Ruiz LM, et al. Copper deficiency alters cell bioenergetics and induces mitochondrial fusion through up-regulation of MFN2 and OPA1 in erythropoietic cells. Biochem Biophys Res Commun. 2013;437(3):426-432. (PubMed)
22. Peled T, Landau E, Prus E, Treves AJ, Nagler A, Fibach E. Cellular copper content modulates differentiation and self-renewal in cultures of cord blood-derived CD34+ cells. Br J Haematol. 2002;116(3):655-661. (PubMed)
23. Lazarchick J. Update on anemia and neutropenia in copper deficiency. Curr Opin Hematol. 2012;19(1):58-60. (PubMed)
24. Shaw JC. Copper deficiency and non-accidental injury. Arch Dis Child. 1988;63(4):448-455. (PubMed)
25. Blackmer AB, Bailey E. Management of copper deficiency in cholestatic infants: review of the literature and a case series. Nutr Clin Pract. 2013;28(1):75-86. (PubMed)
26. Best K, McCoy K, Gemma S, Disilvestro RA. Copper enzyme activities in cystic fibrosis before and after copper supplementation plus or minus zinc. Metabolism. 2004;53(1):37-41. (PubMed)
27. Rowin J, Lewis SL. Copper deficiency myeloneuropathy and pancytopenia secondary to overuse of zinc supplementation. J Neurol Neurosurg Psychiatry. 2005;76(5):750-751. (PubMed)
28. Nations SP, Boyer PJ, Love LA, et al. Denture cream: an unusual source of excess zinc, leading to hypocupremia and neurologic disease. Neurology. 2008;71(9):639-643. (PubMed)
29. Prodan CI, Bottomley SS, Holland NR, Lind SE. Relapsing hypocupraemic myelopathy requiring high-dose oral copper replacement. J Neurol Neurosurg Psychiatry. 2006;77(9):1092-1093. (PubMed)
30. Kumar N, Gross JB, Jr. Mutation in the ATP7A gene may not be responsible for hypocupraemia in copper deficiency myelopathy. Postgrad Med J. 2006;82(968):416. (PubMed)
31. Tumer Z. An overview and update of ATP7A mutations leading to Menkes disease and occipital horn syndrome. Hum Mutat. 2013;34(3):417-429. (PubMed)
32. Kodama H, Fujisawa C, Bhadhprasit W. Inherited copper transport disorders: biochemical mechanisms, diagnosis, and treatment. Curr Drug Metab. 2012;13(3):237-250. (PubMed)
33. Fox PL, Mazumder B, Ehrenwald E, Mukhopadhyay CK. Ceruloplasmin and cardiovascular disease. Free Radic Biol Med. 2000;28(12):1735-1744. (PubMed)
34. Jones AA, DiSilvestro RA, Coleman M, Wagner TL. Copper supplementation of adult men: effects on blood copper enzyme activities and indicators of cardiovascular disease risk. Metabolism. 1997;46(12):1380-1383. (PubMed)
35. Ford ES. Serum copper concentration and coronary heart disease among US adults. Am J Epidemiol. 2000;151(12):1182-1188. (PubMed)
36. Malek F, Jiresova E, Dohnalova A, Koprivova H, Spacek R. Serum copper as a marker of inflammation in prediction of short term outcome in high risk patients with chronic heart failure. Int J Cardiol. 2006;113(2):e51-53. (PubMed)
37. Leone N, Courbon D, Ducimetiere P, Zureik M. Zinc, copper, and magnesium and risks for all-cause, cancer, and cardiovascular mortality. Epidemiology. 2006;17(3):308-314. (PubMed)
38. Kosar F, Sahin I, Acikgoz N, Aksoy Y, Kucukbay Z, Cehreli S. Significance of serum trace element status in patients with rheumatic heart disease: a prospective study. Biol Trace Elem Res. 2005;107(1):1-10. (PubMed)
39. Bertinato J, Zouzoulas A. Considerations in the development of biomarkers of copper status. J AOAC Int. 2009;92(5):1541-1550. (PubMed)
40. Klevay LM. Cardiovascular disease from copper deficiency--a history. J Nutr. 2000;130(2S Suppl):489S-492S. (PubMed)
41. Mielcarz G, Howard AN, Mielcarz B, et al. Leucocyte copper, a marker of copper body status is low in coronary artery disease. J Trace Elem Med Biol. 2001;15(1):31-35. (PubMed)
42. Kinsman GD, Howard AN, Stone DL, Mullins PA. Studies in copper status and atherosclerosis. Biochem Soc Trans. 1990;18(6):1186-1188. (PubMed)
43. Wang XL, Adachi T, Sim AS, Wilcken DE. Plasma extracellular superoxide dismutase levels in an Australian population with coronary artery disease. Arterioscler Thromb Vasc Biol. 1998;18(12):1915-1921. (PubMed)
44. Klevay LM. Lack of a recommended dietary allowance for copper may be hazardous to your health. J Am Coll Nutr. 1998;17(4):322-326. (PubMed)
45. Milne DB, Nielsen FH. Effects of a diet low in copper on copper-status indicators in postmenopausal women. Am J Clin Nutr. 1996;63(3):358-364. (PubMed)
46. Medeiros DM, Milton A, Brunett E, Stacy L. Copper supplementation effects on indicators of copper status and serum cholesterol in adult males. Biol Trace Elem Res. 1991;30(1):19-35. (PubMed)
47. DiSilvestro RA, Joseph EL, Zhang W, Raimo AE, Kim YM. A randomized trial of copper supplementation effects on blood copper enzyme activities and parameters related to cardiovascular health. Metabolism. 2012;61(9):1242-1246. (PubMed)
48. Turley E, McKeown A, Bonham MP, et al. Copper supplementation in humans does not affect the susceptibility of low density lipoprotein to in vitro induced oxidation (FOODCUE project). Free Radic Biol Med. 2000;29(11):1129-1134. (PubMed)
49. Rock E, Mazur A, O'Connor J M, Bonham MP, Rayssiguier Y, Strain JJ. The effect of copper supplementation on red blood cell oxidizability and plasma antioxidants in middle-aged healthy volunteers. Free Radic Biol Med. 2000;28(3):324-329. (PubMed)
50. Mansoor MA, Bergmark C, Haswell SJ, et al. Correlation between plasma total homocysteine and copper in patients with peripheral vascular disease. Clin Chem. 2000;46(3):385-391. (PubMed)
51. Celik C, Bastu E, Abali R, et al. The relationship between copper, homocysteine and early vascular disease in lean women with polycystic ovary syndrome. Gynecol Endocrinol. 2013;29(5):488-491. (PubMed)
52. Gerhard GT, Duell PB. Homocysteine and atherosclerosis. Curr Opin Lipidol. 1999;10(5):417-428. (PubMed)
53. Emsley AM, Jeremy JY, Gomes GN, Angelini GD, Plane F. Investigation of the inhibitory effects of homocysteine and copper on nitric oxide-mediated relaxation of rat isolated aorta. Br J Pharmacol. 1999;126(4):1034-1040. (PubMed)
54. Shukla N, Angelini GD, Jeremy JY. Interactive effects of homocysteine and copper on angiogenesis in porcine isolated saphenous vein. Ann Thorac Surg. 2007;84(1):43-49. (PubMed)
55. Uthus EO, Reeves PG, Saari JT. Copper deficiency decreases plasma homocysteine in rats. J Nutr. 2007;137(6):1370-1374. (PubMed)
56. Wei H, Zhang WJ, McMillen TS, Leboeuf RC, Frei B. Copper chelation by tetrathiomolybdate inhibits vascular inflammation and atherosclerotic lesion development in apolipoprotein E-deficient mice. Atherosclerosis. 2012;223(2):306-313. (PubMed)
57. Failla ML, Hopkins RG. Is low copper status immunosuppressive? Nutr Rev. 1998;56(1 Pt 2):S59-64.
58. Percival SS. Copper and immunity. Am J Clin Nutr. 1998;67(5 Suppl):1064S-1068S. (PubMed)
59. Heresi G, Castillo-Duran C, Munoz C, Arevalo M, Schlesinger L. Phagocytosis and immunoglobulin levels in hypocupremic children. Nutr Res. 1985;5:1327-1334.
60. Kelley DS, Daudu PA, Taylor PC, Mackey BE, Turnlund JR. Effects of low-copper diets on human immune response. Am J Clin Nutr. 1995;62(2):412-416. (PubMed)
61. Hodgkinson V, Petris MJ. Copper homeostasis at the host-pathogen interface. J Biol Chem. 2012;287(17):13549-13555. (PubMed)
62. Looker AC, Melton LJ, 3rd, Harris TB, Borrud LG, Shepherd JA. Prevalence and trends in low femur bone density among older US adults: NHANES 2005-2006 compared with NHANES III. J Bone Miner Res. 2010;25(1):64-71. (PubMed)
63. Tiidus PM, Lowe DA, Brown M. Estrogen replacement and skeletal muscle: mechanisms and population health. J Appl Physiol. 2013;115(5):569-578. (PubMed)
64. Cauley JA. Public Health Impact of Osteoporosis. J Gerontol A Biol Sci Med Sci. 2013;68(10):1243-1251. (PubMed)
65. Kanumakala S, Boneh A, Zacharin M. Pamidronate treatment improves bone mineral density in children with Menkes disease. J Inherit Metab Dis. 2002;25(5):391-398. (PubMed)
66. Marquardt ML, Done SL, Sandrock M, Berdon WE, Feldman KW. Copper deficiency presenting as metabolic bone disease in extremely low birth weight, short-gut infants. Pediatrics. 2012;130(3):e695-698. (PubMed)
67. Baker A, Harvey L, Majask-Newman G, Fairweather-Tait S, Flynn A, Cashman K. Effect of dietary copper intakes on biochemical markers of bone metabolism in healthy adult males. Eur J Clin Nutr. 1999;53(5):408-412. (PubMed)
68. Baker A, Turley E, Bonham MP, et al. No effect of copper supplementation on biochemical markers of bone metabolism in healthy adults. Br J Nutr. 1999;82(4):283-290. (PubMed)
69. Cashman KD, Baker A, Ginty F, et al. No effect of copper supplementation on biochemical markers of bone metabolism in healthy young adult females despite apparently improved copper status. Eur J Clin Nutr. 2001;55(7):525-531. (PubMed)
70. Conlan D, Korula R, Tallentire D. Serum copper levels in elderly patients with femoral-neck fractures. Age Ageing. 1990;19(3):212-214. (PubMed)
71. Mutlu M, Argun M, Kilic E, Saraymen R, Yazar S. Magnesium, zinc and copper status in osteoporotic, osteopenic and normal post-menopausal women. J Int Med Res. 2007;35(5):692-695. (PubMed)
72. Eaton-Evans J, Mellwrath EM, Jackson WE, McCartney H, Strain JJ. Copper supplementation and the maintenance of bone mineral density in middle-aged women. J Trace Elem Exp Med. 1996;9:87-94.
73. Strause L, Saltman P, Smith KT, Bracker M, Andon MB. Spinal bone loss in postmenopausal women supplemented with calcium and trace minerals. J Nutr. 1994;124(7):1060-1064. (PubMed)
74. Nielsen FH, Lukaski HC, Johnson LK, Roughead ZK. Reported zinc, but not copper, intakes influence whole-body bone density, mineral content and T score responses to zinc and copper supplementation in healthy postmenopausal women. Br J Nutr. 2011;106(12):1872-1879. (PubMed)
75. Sidiropoulou-Chatzigiannis S, Kourtidou M, Tsalikis L. The effect of osteoporosis on periodontal status, alveolar bone and orthodontic tooth movement. A literature review. J Int Acad Periodontol. 2007;9(3):77-84. (PubMed)
76. Darcey J, Horner K, Walsh T, Southern H, Marjanovic EJ, Devlin H. Tooth loss and osteoporosis: to assess the association between osteoporosis status and tooth number. Br Dent J. 2013;214(4):E10. (PubMed)
77. Sierpinska T, Konstantynowicz J, Orywal K, Golebiewska M, Szmitkowski M. Copper deficit as a potential pathogenic factor of reduced bone mineral density and severe tooth wear. Osteoporos Int. 2013 [Epub ahead of print]. (PubMed)
78. Squitti R, Barbati G, Rossi L, et al. Excess of nonceruloplasmin serum copper in AD correlates with MMSE, CSF [β]-amyloid, and h-τ. Neurology. 2006;67(1):76-82. (PubMed)
79. Arnal N, Cristalli DO, de Alaniz MJ, Marra CA. Clinical utility of copper, ceruloplasmin, and metallothionein plasma determinations in human neurodegenerative patients and their first-degree relatives. Brain Res. 2010;1319:118-130. (PubMed)
80. Ventriglia M, Bucossi S, Panetta V, Squitti R. Copper in Alzheimer's disease: a meta-analysis of serum, plasma, and cerebrospinal fluid studies. J Alzheimers Dis. 2012;30(4):981-984. (PubMed)
81. Brewer GJ. Copper excess, zinc deficiency, and cognition loss in Alzheimer's disease. Biofactors. 2012;38(2):107-113. (PubMed)
82. Squitti R, Polimanti R. Copper phenotype in Alzheimer's disease: dissecting the pathway. Am J Neurodegener Dis. 2013;2(2):46-56. (PubMed)
83. Squitti R, Polimanti R. Copper hypothesis in the missing hereditability of sporadic Alzheimer's disease: ATP7B gene as potential harbor of rare variants. J Alzheimers Dis. 2012;29(3):493-501. (PubMed)
84. Sparks DL, Schreurs BG. Trace amounts of copper in water induce β-amyloid plaques and learning deficits in a rabbit model of Alzheimer's disease. Proc Natl Acad Sci U S A. 2003;100(19):11065-11069. (PubMed)
85. Kitazawa M, Cheng D, Laferla FM. Chronic copper exposure exacerbates both amyloid and tau pathology and selectively dysregulates cdk5 in a mouse model of AD. J Neurochem. 2009;108(6):1550-1560. (PubMed)
86. Morris MC, Evans DA, Tangney CC, et al. Dietary copper and high saturated and trans fat intakes associated with cognitive decline. Arch Neurol. 2006;63(8):1085-1088. (PubMed)
87. Kessler H, Pajonk FG, Bach D, et al. Effect of copper intake on CSF parameters in patients with mild Alzheimer's disease: a pilot phase 2 clinical trial. J Neural Transm. 2008;115(12):1651-1659. (PubMed)
88. Kessler H, Bayer TA, Bach D, et al. Intake of copper has no effect on cognition in patients with mild Alzheimer's disease: a pilot phase 2 clinical trial. J Neural Transm. 2008;115(8):1181-1187. (PubMed)
89. Skjorringe T, Moller LB, Moos T. Impairment of interrelated iron- and copper homeostatic mechanisms in brain contributes to the pathogenesis of neurodegenerative disorders. Front Pharmacol. 2012;3:169. (PubMed)
90. Akatsu H, Hori A, Yamamoto T, et al. Transition metal abnormalities in progressive dementias. Biometals. 2012;25(2):337-350. (PubMed)
91. Mariani S, Ventriglia M, Simonelli I, et al. Fe and Cu do not differ in Parkinson's disease: a replication study plus meta-analysis. Neurobiol Aging. 2013;34(2):632-633. (PubMed)
92. Hendler SS, Rorvik DR, eds. PDR for Nutritional Supplements. Montvale: Medical Economics Company, Inc; 2001.
93. Bremner I. Manifestations of copper excess. Am J Clin Nutr. 1998;67(5 Suppl):1069S-1073S. (PubMed)
94. Fitzgerald DJ. Safety guidelines for copper in water. Am J Clin Nutr. 1998;67(5 Suppl):1098S-1102S. (PubMed)
95. Turnlund JR, Jacob RA, Keen CL, et al. Long-term high copper intake: effects on indexes of copper status, antioxidant status, and immune function in young men. Am J Clin Nutr. 2004;79(6):1037-1044. (PubMed)
96. Turnlund JR, Keyes WR, Kim SK, Domek JM. Long-term high copper intake: effects on copper absorption, retention, and homeostasis in men. Am J Clin Nutr. 2005;81(4):822-828. (PubMed)
97. Wood RJ, Suter PM, Russell RM. Mineral requirements of elderly people. Am J Clin Nutr. 1995;62(3):493-505. (PubMed)