Contents
Five glutathione peroxidases, three thioredoxin reductases, three iodothyronine deiodinases, and one methionine sulfoxide reductase B1 are among the best characterized selenoproteins with known functions. (More information)
Impaired antioxidant protection in selenium-deficient individuals may affect physiological responses to stress. Keshan cardiomyopathy and Kashin-Beck osteoarthropathy are diseases occurring specifically in areas of selenium deficiency in Asia. (More information)
The current recommended dietary allowance (RDA) set by the US National Academy of Medicine is 55 μg/day for adolescents and most adults. (More information)
Overall, early observational studies have found either null or inverse (protective) associations between selenium exposure and risk of site-specific cancers. However, most recent evidence from intervention trials in selenium-replete participants does not support a protective effect of selenium supplementation against cancer. (More information)
Preliminary evidence from randomized controlled clinical trials suggests that selenium supplementation may prevent viral load progression and increase immune cell count in HIV-positive patients. (More information)
The levels and chemical forms of selenium in plant-based foods vary according to the composition and selenium content of soil in which the plants are grown. Selenium-rich food sources include Brazil nuts (nuts from the Bertholletia excelsa tree), grains, seafood, organ meats, poultry, and dairy products. (More information)
The tolerable upper intake level (UL) for selenium is 400 μg/day for adolescents and adults and includes both selenium obtained from food, which averages about 100 μg/day for adults in the US, and selenium from supplements. (More information)
Because some evidence suggests that high serum selenium concentrations may have adverse effects on glycemic control, individuals with high selenium status and/or those at risk for type 2 diabetes mellitus should avoid taking selenium supplements. (More information)
Selenium is a trace mineral that is essential for humans in small amounts, but like all essential elements, selenium can be toxic at high levels. Unlike plants, most animals — including humans — require selenium for the appropriate functioning of a number of selenium-dependent enzymes known as selenoproteins. During protein synthesis (translation), the amino acid selenocysteine is incorporated into elongating proteins at very specific locations in the amino acid sequence in order to form functional selenoproteins. Although higher plants do not appear to require selenium for survival, they can incorporate it nonspecifically into sulfur-containing molecules when the mineral is present in the soil (1). Of note, in animals, the amino acid selenomethionine can be nonspecifically incorporated into proteins in place of methionine (2). However, only selenocysteine-containing proteins are regarded as selenoproteins (Figure 1).

Twenty-five genes coding for selenoproteins have been identified in humans (3). The insertion of selenocysteine into selenoproteins during translation is directed by the presence of a selenocysteine-insertion sequence (SECIS) within selenoprotein mRNAs. Briefly, the recognition of SECIS by the translational machinery results in the recruitment of specific translational factors that decode in-frame UGA codons by inserting selenocysteine into elongating selenoproteins (4).
Research is gradually uncovering the metabolic functions of all human selenoproteins, including splicing variants (5). Some of the selenoproteins with identified functions are discussed below.
Five selenium-containing glutathione peroxidases (GPx1-4 and GPx6) have been identified: GPx1 (cytosolic GPx), GPx2 (epithelial cell-specific GPx first identified in intestinal lining and lungs), GPx3 (highly expressed in thyroid gland and kidneys), GPx4 (phospholipid-hydroperoxide GPx; PHGPx), and GPx6 (expressed in the olfactory epithelium) (6). GPx isoenzymes are all antioxidant enzymes that reduce potentially damaging reactive oxygen species (ROS), such as hydrogen peroxide and lipid hydroperoxides, to harmless products like water and alcohols by coupling their reduction with the oxidation of glutathione (Figure 2a). Spermatogenesis and male fertility are highly dependent on GPx4 and selenoprotein P (SELENOP, formerly SEPP1 or SELP; see below). In the testes, GPx4 reduces phospholipid hydroperoxides, hence protecting immature spermatozoa cells against oxidative stress. GPx4 is also a major structural protein of the capsule embedding mature sperm mitochondrial helix involved in sperm motility. SELENOP is essential for selenium supply to the testes, and animal models lacking the SELENOP gene are infertile due to poor selenium tissue bioavailability, defective GPx4 synthesis, and impaired sperm maturation (7).

In mammals, three selenocysteine-containing thioredoxin reductase (TXNRD) isoenzymes have been identified in the thioredoxin system: cytosolic TXNRD1, mitochondrial TXNRD3, and testes-specific thioredoxin glutathione reductase (TXNRD3, also known as TGR). TXNRDs are homodimeric enzymes, and each monomer contains FAD- and NADPH-binding domains and a selenocysteine-containing catalytic site. TXNRDs catalyze the reduction of a wide range of substrates, including thioredoxin and protein disulfide isomerase (PDI) (see Figure 2b). TXNRDs also serve as electron donors for the regeneration of small antioxidants, possibly recycling ascorbic acid (vitamin C), α-lipoic acid, α-tocopherol (vitamin E), and coenzyme Q10 from their oxidized forms (8). The maintenance of thioredoxin in a reduced form by TXNRDs is important for regulating cell growth and survival. The protein thioredoxin, together with TXNRD1 (or TXNRD3), NADPH, and FAD, constitute the thioredoxin antioxidant system involved in the reduction of antioxidant enzymes (e.g., peroxiredoxins, methionine sulfoxide reductases, and ribonucleotide reductase) and of many oxidation/reduction (redox)-sensitive signaling proteins (9). TXNRD1 is one of the most investigated selenoproteins and regarded as one of the major antioxidant enzymes and redox regulators in mammalian cells.
The thyroid gland releases very small amounts of biologically active thyroid hormone (triiodothyronine or T3) and larger amounts of an inactive form of thyroid hormone (T3 precursor: thyroxine or T4) into the circulation. Most of the biologically active T3 in the circulation and inside cells is generated by the removal of one iodine atom from T4 in a reaction catalyzed by selenium-dependent iodothyronine deiodinase enzymes. Two different selenium-dependent iodothyronine deiodinases (DIOs type 1 and 2) can deiodinate T4, thus increasing circulating T3, while a third iodothyronine deiodinase (DIO type 3) can convert both T3 and T4 to inactive metabolites (Figure 3) (10, 11). Of note, inactivation of the genes encoding DIOs in rodent models has revealed a role for DIO type 1 in iodine homeostasis and the importance of DIOs type 2 and 3 in the maturation of auditory and visual systems during fetal development (10). Thus the importance of selenium in normal development, growth, and metabolism is not limited to its role in the regulation of thyroid gland function.

Selenoprotein P (SELENOP, formerly SEPP1 or SELP) is predominantly produced by the liver, a major storage site for selenium, and secreted in the plasma. The full-length glycoprotein contains a selenium-rich domain with nine selenocysteine residues, as well as a thioredoxin-like catalytic site with one selenocysteine residue. SELENOP constitutes the major form of selenium transport to peripheral tissues (12, 13). SELENOP also functions as an antioxidant that protects cells from oxidative damage by enabling full activity of thioredoxin reductases and glutathione peroxidases through adequate supply of selenium to extrahepatic tissues (see Glutathione peroxidases). SELENOP appears to be especially critical for selenium homeostasis in the brain and testes where apolipoprotein E receptor 2 (apoER2) facilitates the uptake of SELENOP. Megalin is another SELENOP-specific lipoprotein receptor that helps limit urinary selenium loss through SEPP1 reuptake by the kidneys (3, 14). SEPP1 has been implicated in the regulation of glucose metabolism and insulin sensitivity (15). Moreover, low plasma concentrations of SELENOP have been associated with increased risk of heart failure in a recent population-based, prospective cohort study (16).
Selenoprotein W (SELENOW, formerly SEPW or SelW) exists in different isoforms (homologues) and is highly conserved across species. In humans, SELENOW is expressed in numerous tissues, with highest levels found in skeletal muscle and heart (17). SELENOW contains a selenocysteine residue and a cysteine residue that binds to a glutathione molecule, suggesting a role in redox regulation (18). The expression of SELENOW is correlated with selenium status and appears to be sensitive to low-selenium supply (19, 20). SELENOW expression in the brain of mice has been found to confer protection against oxidative stress-induced neuronal cell death (21). SELENOW also appears to be a negative regulator for 14-3-3 proteins. Indeed, 14-3-3 inhibition by SELENOW in breast cancer cells was found to increase cell proliferation and cell survival through increasing resistance to genotoxic stress in human breast and lung cancer cells (22). In skeletal muscle cells, SELENOW was shown to reduce the binding of 14-3-3 to a transcriptional co-activator with PDZ-binding motif (TAZ) (23), allowing TAZ translocation to the nucleus and subsequent activation of muscle cell differentiation genes (24). Finally, SELENOW was found to prevent the degradation of the epidermal growth factor receptor (EGFR) in breast and prostate epithelial cells in culture. EGFR is constitutively activated in many tumors, and evidence of a role for SELENOW in EGFR activation and signaling may help shed light on the relationship between selenium status and cancer risk (25).
There is no free pool of the amino acid selenocysteine in cells such that selenocysteine synthesis takes place on a specialized tRNA immediately preceding the translation of selenoprotein mRNAs. The reaction is catalyzed by pyridoxal 5’-phosphate (PLP)-dependent L-seryl-tRNASec selenium transferase and uses selenophosphate (monoselenium phosphate) as the selenium donor (Figure 4) (26). Selenophosphate synthetase 2 is the selenoenzyme that catalyzes the ATP-dependent synthesis of selenophosphate from hydrogen selenide (Figure 4) (5).

The methionine sulfoxide reduction system is involved in the protection against oxidative stress and is especially critical for the regeneration of proteins damaged by ROS. Indeed, ROS can oxidize methionine residues (methionine sulfoxides) within proteins and potentially impair their activities. In humans, two stereospecific families of methionine sulfoxide reductases (MsrA and MsrB) are encoded by a single MSRA gene and three MSRB genes (MSRB1-3). MsrA catalyzes the reduction of the S-form of methionine sulfoxide; the R-form of methionine sulfoxide is reduced by MsrB1, 2, or 3. Only MsrB1 has been characterized as a selenoprotein with one selenocysteine residue in its catalytic site. MsrB1 appears to be involved in the redox regulation of certain proteins. In macrophages, reorganization of the actin cytoskeleton necessary for chemotaxis and phagocytosis requires MsrB1-dependent reduction of methionine-R-sulfoxide residues within actin (27). Studies using MSR gene inactivation in mice have also shown that methionine sulfoxide reduction is implicated in the regulation of the methionine cycle (reviewed in 28). Both the thioredoxin (Trx) and GSH-dependent glutaredoxin (Grx) antioxidant systems have been found to reduce methionine sulfoxide reductases in vitro and/or in vivo (see Figure 2) (28).
The 15 kDa selenoprotein SELENOF (formerly selenoprotein 15; SEP15) is highly expressed in several tissues, including prostate, kidney, testes, liver, and brain (29). Although its function continues to be elucidated, SELENOF was found to interact with the endoplasmic reticulum UDP-glucose:glycoprotein glucosyltransferase (UGGT), an enzyme involved in the quality control of glycoprotein folding in the endoplasmic reticulum (30, 31). Because SELENOF has a thioredoxin-like catalytic site, SELENOF is thought to either regulate UGGT activity or the redox state of UGGT substrates (32). Mice lacking a functional SELENOF were found to develop nuclear cataract (lens opacification) at a very early age suggesting that SELENOF may be critical to the quality control system of protein folding in the lens (33). SELENOF may also be implicated in tissue-specific anticancer mechanisms (reviewed in 34), including colorectal cancer, that may, in part, be the result of SELENOFs potential role in intestinal barrier integrity (35).
The mammalian selenoprotein S (SELENOS, formerly known as SEPS1, SelS, or VCP-interacting membrane selenoprotein [VIMP]) is another endoplasmic reticulum (ER) membrane protein. SELENOS is involved in the cellular response to ER stress (ER-associated degradation; ERAD) activated by the detection of misfolded proteins. SEPS1 contributes to the removal and transfer (retrotranslocation) of misfolded proteins from the ER lumen to the cytosol where proteins are tagged with ubiquitin before being degraded. A polymorphism or variation in the sequence within an ER-response element located in the SELENOS promoter was found to result in reduced SELENOS promoter activity and gene expression (36). The polymorphism corresponding to the substitution of a guanine (G) by an adenine (A) at nucleotide -105 (-105G>A) has been associated with increased plasma concentrations of pro-inflammatory cytokines. In addition, a case-control study reported that the A allele was more prevalent in individuals affected by Hashimoto thyroiditis (HT) — a T-cell-mediated autoimmune disease resulting in the destruction of thyroid cells — than in healthy controls (37). Other associations between SELENOS polymorphism (including -105G>A) and susceptibility to various conditions, such as preeclampsia, coronary artery disease, or gastrointestinal cancers, strongly suggest a role for this selenoprotein in the regulation of inflammatory and immune responses (38-41).
Other less well-characterized selenoproteins, which are also localized in the ER lumen and/or membrane, include selenoproteins K (SELENOK), M (SELENOM), N (SELENON), and T (SELENOT) (42).
The importance of selenium to biological systems, and specifically to the cellular redox (pro-oxidant/antioxidant) balance, is thought to be derived primarily from its presence as selenocysteine in the catalytic site of selenoproteins (see Function). Other minerals that are critical components of antioxidant enzymes include copper (as superoxide dismutase), zinc (as superoxide dismutase), and iron (as catalase). Selenium acts via selenoenzymes in synergy with the antioxidant vitamins, vitamin C (ascorbic acid) and vitamin E (α-tocopherol), by regenerating them from their oxidized forms and promoting maximal antioxidant protection (43-45).
While iodine is an essential component of thyroid hormones, the selenium-containing iodothyronine deiodinases (DIOs) are enzymes required for the conversion of thyroxine (T4) to the biologically active thyroid hormone, triiodothyronine (T3) (see Function). DIO1 activity may also be involved in regulating iodine homeostasis (46). Additionally, the glutathione peroxidases play a critical role in thyroid function because they catalyze the degradation of peroxides generated during thyroid hormone synthesis (10). The epidemiology of coexisting iodine and selenium deficiencies in central Africa, but not in China, has been linked to myxedematous cretinism, a severe form of congenital hypothyroidism accompanied by mental and physical retardation. Selenium deficiency may be only one of several undetermined factors that might exacerbate the detrimental effects of iodine deficiency (47). Interestingly, selenium deficiency in rodents was found to have little impact on DIO activities as it appears that selenium is being supplied in priority for adequate synthesis of DIOs at the expense of other selenoenzymes (10).
Insufficient selenium intake may negatively affect the activity of several selenium-responsive enzymes, including glutathione peroxidases (specifically, GPx1 and GPx3), iodothyronine deiodinases, selenoprotein W, and methionine-R-sulfoxide reductase B1 (MsrB1). Even when severe, isolated selenium deficiency does not usually result in obvious clinical illness. Yet, compared to subjects with adequate selenium status, selenium-deficient individuals might be more susceptible to additional physiological stresses (48). Prolonged selenium deficiency may likely contribute to Keshan and Kashin-Beck diseases (see below).
Selenium deficiency has been reported in chronically ill patients who received total parenteral nutrition (TPN) without added selenium for prolonged periods of time. Muscular weakness, muscle wasting, and cardiomyopathy (inflammation and damage to the heart muscle) have been observed in these patients. Today, TPN solutions are routinely supplemented with selenium.
The risk of selenium deficiency also may be increased following bariatric surgery or in severe gastrointestinal conditions, such as Crohn's disease (49, 50). Some specialized medical diets like those used to treat certain metabolic disorders, including phenylketonuria, homocystinuria, and maple syrup urine disease, need to be supplemented with selenium to ensure optimal selenium status in patients (51).
Keshan disease is a fatal form of dilated cardiomyopathy that was first described in young women and children in a selenium-deficient region in China. The acute form of the disease is characterized by the sudden onset of cardiac insufficiency, while the chronic form results in moderate-to-severe heart enlargement with varying degrees of cardiac insufficiency (52). The incidence of Keshan disease is closely associated with very low dietary intakes of selenium and poor selenium nutritional status. Selenium supplementation (in the form of sodium selenite; Na2SeO3) was found to protect people from developing Keshan disease but could not reverse heart muscle damage once it had occurred (52). One case-control study reported that selenium-responsive glutathione peroxidase 1 (GPx1) activity was significantly lower in Keshan patients compared to healthy individuals. Interestingly, a specific GPX1 polymorphism resulting in a proline-to-leucine transition at position 198 (Pro198Leu) is associated with a reduction in GPx1 activity and found to be more prevalent in Keshan patients. This GPX1 polymorphism might confer a greater susceptibility to Keshan disease in carriers with low selenium nutritional status (53).
While selenium deficiency is a major etiological factor of Keshan disease (54), the seasonal and annual variation in disease occurrence suggested that other factors, especially an infectious agent, might be involved in addition to selenium deficiency (55). Coxsackie virus B3 is one virus type that has been isolated from Keshan patients, and animal studies have shown that this virus was capable of causing an inflammation of the heart (myocarditis) in selenium-deficient mice. Studies in mice also indicated that oxidative stress induced by selenium deficiency could result in changes in the viral genome, such as to convert a relatively harmless strain of coxsackie virus B3 into a myocarditis-causing strain (56). Although not documented in Keshan disease, it is possible that selenium deficiency may increase the virulence of viruses with the potential to invade and damage the heart muscle (57).
Kashin-Beck disease (KBD) is an endemic condition that affects an estimated 2.5 million people in Tibet, northern and central China, North Korea, and southeastern Siberia, a region of Russia (58). KBD is characterized by the degeneration of articular cartilage between joints (osteoarthritis) that can result in joint deformities and dwarfism in the most severe forms of the disease. The disease affects children as young as two years old. As with Keshan disease, KBD is prevalent in selenium-deficient provinces and thus generally affects people with very low selenium intakes (58, 59). Studies have suggested that increased susceptibility to KBD in selenium-deficient populations might result from a reduced antioxidant protection associated with polymorphisms in GPX genes (60, 61), but polymorphisms in other genes have also been implicated (62). Yet, the etiology appears to be multifactorial, as a number of other causative factors have been suggested for KBD, including fungal toxins in grain, iodine deficiency, and contaminated drinking water (52).
Meta-analyses of a few small clinical trials and prospective cohort studies have indicated that improving selenium nutritional status in children living in endemic areas may help reduce KBD incidence (63). Also, there is limited evidence to suggest that selenium supplementation could be useful in the treatment of patients with KBD. A meta-analysis of 10 randomized controlled trials reported a significant increase in the repairing rate of bone lesions in KBD children supplemented with sodium selenite for at least one year (64). Larger trials of higher quality are needed to assess whether selenium supplementation could result in disease remission.
The dietary reference intakes (DRIs) for selenium were last revised in 2000 by the Food and Nutrition Board of the US Institute of Medicine (now the National Academy of Medicine). The most recent RDA (Table 1) is based on the estimated average requirement (EAR) needed to maximize antioxidant enzyme glutathione peroxidase (GPx) activity in plasma (65).
| Life Stage | Age | Males (μg/day) | Females (μg/day) |
|---|---|---|---|
| Infants | 0-6 months | 15 (AI) | 15 (AI) |
| Infants | 7-12 months | 20 (AI) | 20 (AI) |
| Children | 1-3 years | 20 | 20 |
| Children | 4-8 years | 30 | 30 |
| Children | 9-13 years | 40 | 40 |
| Adolescents | 14-18 years | 55 | 55 |
| Adults | 19 years and older | 55 | 55 |
| Pregnancy | all ages | - | 60 |
| Breast-feeding | all ages | - | 70 |
Of note, a US national survey (NHANES III) reported that over 99% of the US participants had serum selenium concentrations consistent with selenium requirements being met (65), suggesting selenium supplementation is not needed for most Americans.
There has been considerable research on the effect of selenium supplementation on the incidence of cancer using preclinical models. More than two-thirds of over 100 published studies in 20 different animal models of spontaneous, viral, and chemically induced cancers found that selenium supplementation (to at least adequate intake levels) significantly reduces tumor incidence, in comparison to selenium-deficient diets (66). Evidence of cancer-inhibiting effects of selenium has provided a strong rationale for investigating potential associations between selenium intake and cancer risk in humans.
Most of the early observational evidence from case-control and nested case-control studies suggested either null or inverse associations between selenium exposure and risk of site-specific cancers (67). Biomarkers of selenium exposure include toenail and blood selenium content, as well as plasma glutathione peroxidase (GPx) activity. However, it is not clear whether these biomarkers adequately reflect selenium exposure from dietary and supplemental sources or selenium distribution in tissues and organs that may be affected by cancer. In the Danish Prospective Diet, Cancer, and Health prospective study that followed over 57,000 men and women for 14 years, the risk of rectal cancer was found to be 42% higher in current smokers compared to nonsmokers. No difference between smokers and nonsmokers regarding supplemental and dietary intakes of antioxidant micronutrients, including selenium, was found to contribute to the association of smoking and rectal cancer (68). Yet, because studies have consistently reported lower blood selenium concentrations and GPx activities in smokers compared to nonsmokers (reviewed in 69), estimation of selenium intakes might not be a reliable marker of selenium exposure in this population. Also, the chemical forms of selenium found in food are varied (see Sources) and may have very different biological and toxicological effects (70, 71).
A Cochrane review included 55 completed observational studies — mostly with a nested case-control design — published over three decades (67). A meta-analysis of 16 of these observational studies, including over 144,000 individuals, reported that higher versus lower selenium status was associated with a 31% lower risk of cancer at any site and a 40% lower risk of cancer-related mortality. A significantly lower risk was reported for bladder cancer (5 studies) and prostate cancer (17 studies); however, higher selenium status was not inversely associated with risks of breast cancer (8 studies), lung cancer (12 studies), colorectal cancer (5 studies), and gastric cancer (5 studies) (67). Other meta-analyses of observational studies have reported inverse associations between serum selenium concentrations and breast cancer (72) and thyroid cancer (73). Sex differences in cancer susceptibility have been reported in some studies, although consistent evidence of different effects in men and women appears to be lacking.
Single nucleotide variations (polymorphisms) in the sequence of genes can modify gene expression level and the stability and activity of the synthesized proteins. For example, a proline-to-leucine transition caused by a specific polymorphism in the GPX1 gene (rs1050450 C>T) is associated with reduced GPx1 enzymatic activity. The description of several polymorphisms in genes encoding selenoproteins has led to evaluation of possible associations with selenium status and cancer incidence. Notably, certain polymorphisms in genes coding for selenoproteins have been associated with increased risks of gastric and colorectal cancers (74, 75). Additionally, a number of studies have investigated the associations among selenoprotein polymorphisms, selenium status, and prostate cancer risk. A nested case-control study within the EPIC-Heidelberg cohort combined genotyping for several selenoprotein variants with biomarkers of selenium status (76). Briefly, the study showed that a GPX1 gene polymorphism (rs1050450 C>T) affected the association between selenium concentrations and prostate cancer risk. Specifically, selenium concentrations were found to be inversely associated with prostate cancer risk only among carriers of the GPX1 T allele. Additional variants in selenoprotein genes may mitigate the effect of selenium status on the risk of prostate cancer (77, 78). In another nested case-control study within the Physicians’ Health Study (PHS), individuals in the highest versus lowest quartile of plasma selenium concentrations were found to have a reduced risk of prostate cancer-related mortality except if carrying a specific variant in 15kDa selenoprotein gene (SELENOF rs561104 G>A) (79). More research is needed to further unravel the mechanisms underlying the influence of gene-diet interactions on the risk of developing cancer.
Community-based studies: An early intervention trial of selenium supplementation was undertaken in China in 1985 among a general population of 130,471 individuals living in a high-risk area for viral hepatitis B infection and liver cancer. The trial provided table salt enriched with sodium selenite to the population of one township (20,847 people) using four other townships as controls. During an eight-year follow-up period, the average incidence of liver cancer was reduced by 35% in the selenium-supplemented population, while no reduction was found in the control populations. In a clinical trial in the same region in 1987-1990, 226 individuals with evidence of chronic hepatitis B infection were supplemented daily with either 200 μg of selenium in the form of a selenium-enriched yeast tablet or a placebo yeast tablet. During the four-year follow-up period, 7 out of 113 individuals on the placebo developed primary liver cancer, while none of the 113 subjects supplemented with selenium developed liver cancer (80).
Randomized controlled trials: The double-blind, placebo-controlled Nutritional Prevention of Cancer (NPC) study in 1,312 older adults with a history of nonmelanoma skin cancer found that supplementation with 200 μg/day of selenium-enriched yeast (selenized yeast) for an average of 7.4 years resulted in a 52% decrease in prostate cancer incidence in men (81). The protective effect of selenium supplementation was greatest in men with lower baseline plasma selenium and prostate-specific antigen (PSA) levels. A reduced incidence in lung, colorectal, and total cancer was also associated with supplementation with 200 μg/day (82) but not with 400 μg/day of selenium-enriched yeast (83). Additionally, selenium supplementation increased the risk of one type of skin cancer (squamous cell carcinoma) by 25%. A larger randomized, placebo-controlled intervention trial (the SELECT study) in more than 35,000 middle-aged and selenium-replete men, randomized to receive selenium (in the form of selenomethionine, 200 μg/day) and/or vitamin E supplementation, was halted because of concerns regarding an increased risk of type 2 diabetes mellitus with selenium and increased risk of prostate cancer with vitamin E (84, 85). In addition, the supplementation of selenium, alone or together with vitamin E, did not show any benefits regarding the risk of prostate, lung, or colorectal cancers after 5.5 years of follow-up (86, 87). In a randomized, placebo-controlled trial in 1,374 participants who had colonoscopic removal of at least one colorectal adenoma (the Selenium and Celecoxib trial), selenium supplementation (200 μg/day with selenized yeast) for a median of 33 months had no effect on colorectal adenoma recurrence (88). However, in a subanalysis of patients with advanced adenoma at baseline (n=161), selenium supplementation reduced adenoma recurrence by 18% (88). Outcomes from other smaller trials (reviewed in 89) have suggested either a lack of an effect or the possibility of an increased risk of cancer. The lack of a beneficial effect of selenium supplementation was supported in a 2014 meta-analysis of randomized controlled trials (67).
Low activity levels of the selenoenzymes, glutathione peroxidases (GPx), have been reported in oxidative stress-related diseases, including cardiovascular disease (CVD) (90). Presumably, the maintenance of an optimal selenium status has the potential to protect against oxidative stress (including lipid peroxidation) and could possibly aid in the prevention of chronic inflammation and cardiovascular disorders. However, most of the available research to date on selenium status and risk of CVD comes from observational studies, and results are largely conflicting.
Analyses of cross-sectional data from 13,887 US adults included in the Third National Health and Nutrition Examination Survey (NHANES III, 1988-1994) failed to show any significant associations between serum selenium concentrations and mortality from CVD, coronary artery/heart disease (CAD), or stroke (91). In addition, while individuals with renal insufficiency are at higher risk of developing CAD compared to those with normal kidney function, that risk was not found to be greater with low rather than normal selenium concentrations in serum (≤98 μg/L vs. >98 μg/L) (92). Yet, analysis of data from NHANES 2003-2018, a US national cross-sectional survey of 39,438 adults, found an inverse association between selenium intake and stroke, with daily intakes of 105 μg/day linked to the lowest stroke risk (93). A meta-analysis of 12 observational studies (5 prospective cohort studies, 4 case-control studies, and 3 cross-sectional studies) also found blood concentrations of selenium to be inversely associated with risk of stroke (94).
Some observational studies have raised concern with high selenium status. A cross-sectional study based on NHANES 2003-2004 data from 2,638 participants ages 40 years and over found that the risk of high blood pressure (hypertension), a major contributing factor for CVD, was 73% higher in individuals in the upper versus lowest quintile of serum selenium concentrations (≥150 μg/L vs. <122 μg/L) (95). But a systematic review of the literature failed to find enough evidence to support any relationship between serum selenium concentrations and hypertension (96). A few observational studies have also reported associations between normal-to-high selenium status and elevated serum lipid levels in selenium-replete populations, speculating that selenium might interfere with lipid metabolism and adversely affect cardiovascular health (97, 98).
At present, randomized controlled trials have not provided consistent results regarding the effect of selenium supplementation on lipid levels nor have they demonstrated any additional cardiovascular benefits of selenium in individuals with suboptimal or optimal selenium intakes (99, 100). A recent meta-analysis that pooled randomized controlled trials reported no association of single-nutrient selenium supplementation with cardiovascular disease (4 trials), coronary heart disease (3 trials), stroke (3 trials), or cardiovascular-related mortality (5 trials) (101).
A meta-analysis that combined five observational studies and one randomized controlled trial found selenium status was not significantly associated with incidence of cardiovascular disease (RR, 0.66; 95% CI, 0.40-1.09), but the data were highly heterogenous (102). Yet, when 11 studies were included in a dose-response meta-analysis, a 15% lower risk in CVD incidence with each incremental increase of 10 μg/L in blood selenium concentration was found; no association with CVD incidence was seen in a dose-response analysis of eight studies measuring selenium status by toenail concentration (102).
Selenium deficiency has been associated with impaired immunity and chronic inflammation (103). A considerable amount of research conducted in cell culture and animal models indicates that selenium plays essential roles in regulating the migration, proliferation, differentiation, activation, and optimal function of immune cells, thus influencing innate immunity, B-cell dependent antibody production, and T-cell immunity (reviewed in 104). A review of nine randomized controlled trials concluded that selenium supplementation may affect cell-mediated (T-cells, natural killer cells) immunity but has little effect on antibody-mediated (humoral) immunity (105).
Evidence on the role of selenium and selenoproteins in the production of lipid mediators (called eicosanoids) involved in inflammatory responses suggests that selenium supplementation might mitigate dysfunctional inflammatory responses that contribute to the pathogenesis of many chronic health conditions (106). At present, randomized controlled trials are needed to evaluate the potential benefits of selenium supplementation in inflammatory disorders, such as asthma (107) and inflammatory bowel disease (108).
In areas of widespread malnutrition, deficiencies in micronutrients (including selenium) are common among individuals infected by the human immunodeficiency virus (HIV) that causes acquired immunodeficiency syndrome (AIDS). Before antiretroviral therapy (ART) became the standard for HIV treatment, observational studies had consistently reported associations between low serum selenium concentrations and HIV infection in well-nourished subjects (109). Poor selenium status has been linked to increased risks of dilated cardiomyopathy and mortality in HIV-infected children and adults, as well as mother-to-child HIV transmission and perinatal mortality (reviewed in 110). Early laboratory studies suggested that HIV might disrupt normal antioxidant defenses in infected T-cells by reducing the levels of selenoproteins, i.e., thioredoxine reductases and glutathione peroxidases (110). Interestingly, a cross-sectional study found that HIV-seropositive individuals receiving ART for more than two years had undetectable plasma viral loads, higher CD4 lymphocyte T-cell counts, and adequate serum selenium concentrations compared to ART-naïve subjects (111). Because the antioxidant activity of selenoproteins may interfere with viral replication in HIV-infected immune cells (112, 113), it has been suggested that selenium supplementation might serve as a potential adjunct to ART for HIV patients.
A few trials of selenium supplementation in HIV-infected individuals have been conducted. A randomized, double-blind, placebo-controlled trial in 186 HIV-positive adults initially found that selenium supplementation at 200 μg/day for two years significantly decreased the rate of hospital admissions (114). Another randomized, double-blind, placebo-controlled trial in 174 HIV-positive individuals reported that 200 μg/day of selenium supplementation (in the form of selenium-enriched yeast) for nine months increased serum selenium concentrations, improved CD4 lymphocyte T-cell count, and prevented any progression of the HIV viral load (115). In a third double-blind trial in Tanzania, 913 pregnant women between 12 and 27 weeks’ gestation were randomized to receive 200 μg/day of selenium (as selenomethionine) or a placebo until six months after birth. Selenium supplementation had no effect on maternal CD4, CD8, and CD3 T-cell counts and on HIV viral load, but it significantly decreased the risk of acute or persistent diarrhea (116, 117). In addition, the risk of death between six weeks and six months postpartum was significantly reduced in infants of mothers supplemented with selenium compared to placebo (117).
A four-armed trial in Botswana randomized 878 HIV-positive adults at an early stage of the infection to receive either a placebo treatment, multivitamins (vitamins B, C, and E), 200 μg/day of selenium, or both multivitamins and selenium for 24 months (118). Unlike selenium alone, supplementation with multivitamins (with or without selenium) reduced the risk of immune decline by significantly increasing the time before ART initiation became necessary (i.e., when CD4 T-cell count fell below 251 cells/mL) compared to placebo. In the study, a combined outcome of (1) CD4 T-cell count falling below 251 cells/mL; (2) occurrence of AIDS-defining conditions; and (3) AIDS-related death — whichever happened first — was used to evaluate disease progression in the different arms of treatment. Compared to placebo, there was a longer period of time from randomization to the date of the composite outcome in individuals supplemented with multivitamins plus selenium, but not in those who received multivitamins or selenium alone (118). Moreover, a systematic review of six randomized controlled trials found that selenium supplementation (200 μg/day for 9 to 24 months) in HIV-infected subjects did not suppress the virus but delayed the decline in CD4 T cells in most trials (119).
The systemic inflammatory response syndrome (SIRS) results from a systemic inflammatory response that can be due to an infection (sepsis) (120). Severe sepsis and septic shock — defined as persistent sepsis-induced low blood pressure — are associated with elevated mortality rates in critically ill patients (120, 121). Because systemic inflammatory responses involve excessive oxidative stress, it has been suggested that providing antioxidant nutrients like selenium may improve the outcome of critically ill patients in intensive care units. Two meta-analyses of randomized controlled trials found that intravenous selenium supplementation (as sodium selenite) in critically ill patients with SIRS, sepsis, or septic shock resulted in significantly reducing the risk of mortality by 17% to 27% (122, 123). However, in a more recent, large placebo-controlled trial in 538 patients with severe sepsis or septic shock, intravenous selenium selenite administration did not improve mortality at the 28-day mark (124). International guidelines do not support the use of intravenous selenium in the treatment of sepsis and septic shock (125).
Hashimoto thyroiditis (HT; chronic autoimmune thyroiditis) is an autoimmune disease characterized by T-cell infiltration in the thyroid gland and circulating autoantibodies (predominantly against thyroid peroxidase but also against thyroglobulin), causing prolonged inflammation, tissue damage, and hypothyroidism (10) and increased risk of papillary thyroid cancer (126). While the function of the thyroid gland of healthy individuals is usually protected from variations in selenium supply, selenium deficiency and genetic polymorphisms affecting the activity of selenoproteins might be potential contributing factors to autoimmune thyroid diseases. A Cochrane systematic review (127) identified four randomized controlled trials that evaluated the effect of selenium supplementation as an adjunct treatment to T4 replacement therapy (levothyroxine) in HT patients (128-131). While three out of four studies suggested a reduction in levels of circulating autoantibodies, none of them provided information on whether selenium may improve mood- and health-related symptoms to allow for a decreased dosage of levothyroxine. In a recent open-label trial in 90 patients newly diagnosed with HT and not taking levothyroxine, supplementation with 200 μg/day in the form of selenious yeast for six months lowered blood concentrations of autoantibodies (both thyroid peroxidase and thyroglobulin) compared to the ‘no treatment’ group (132). Placebo-controlled studies are needed to evaluate whether supplemental selenium might improve clinical symptoms or influence the dosage of or need for levothyroxine. At present, evidence from randomized controlled trials is largely lacking.
Graves’ disease is an autoimmune thyroid disease that leads to hyperthyroidism. One randomized controlled trial found that selenium supplementation improved the well-being of patients affected by this disease (133). The results of two ongoing, randomized, placebo-controlled trials — the CATALYST in HT patients and the GRASS trial in patients with Graves’ disease — may provide insight into an effect of selenium on thyroid-specific quality-of-life criteria and inform clinical decision making (134, 135).
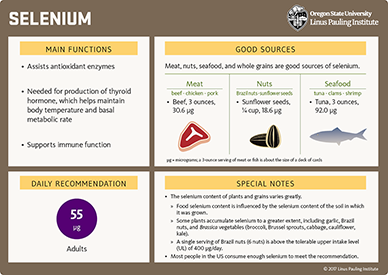
The richest food sources of selenium are organ meats and seafood, followed by muscle meats from farmed animals, as many are supplemented with selenium in their feed. Drinking water is not considered to be a significant source of selenium in North America. However, in areas where high levels of selenium in soil contribute to the selenium content of the water (e.g., California, the Dakotas), higher levels of selenium may be found in wells used for drinking water (136). In general, there is wide variation in the selenium content of plants and grains, especially because some plants, including garlic, Brazil nuts (nuts from the Bertholletia excelsa tree), and multiple Brassica species, tend to accumulate selenium ('selenium accumulators'), while others assimilate selenium to a lesser extent ('non-accumulators'). The assimilation of selenium by plants also depends on soil selenium content. Brazil nuts grown in areas of Brazil with selenium-rich soil may provide more than 100 μg of selenium in one nut, while those grown in selenium-poor soil provide an amount 10 times lower (137). In the US, grains are a good source of selenium, but fruit and vegetables tend to be relatively poor in selenium.
Various chemical forms (species) of selenium are found in selenium accumulators, including selenate (inorganic selenium), selenomethionine, selenocysteine, selenium-methyl-selenocysteine, and γ-glutamyl-selenium-methyl-selenocysteine. Although the two latter compounds are predominant in plants of the Allium and Brassicaceae families (which include garlic, onion, and broccoli), wheat, other grains (including Brazil nuts), and soy are rich in selenomethionine and contain smaller amounts of selenocysteine and selenate. Less is known about selenium species and distribution in dietary sources of animal origin. Animal nutrition and growth conditions certainly contribute to the selenium species formed and their relative quantities, and it is assumed that the metabolic pathway of dietary selenium in animals is similar to that in humans. Selenocysteine is predominantly formed in animals fed inorganic selenium, while selenomethionine is derived from dietary sources of plant origin (reviewed in 138).
In the US, the national survey NHANES III reported mean dietary intakes ranging between 100.5 μg/day and 158.5 μg/day in adults ages 19-50 years (65). Table 2 lists some good food sources of selenium and their average selenium content in micrograms (μg). For more information on the selenium content of specific foods, search USDA’s FoodData Central.
| Food | Serving | Selenium (μg) |
|---|---|---|
| Brazil nuts (from selenium-rich soil) | 1 ounce (6 kernels) | 544* |
| Oysters (Pacific, steamed) | 3 ounces | 131 |
| Tuna (yellowfin, cooked, dry heat) | 3 ounces | 91.8 |
| Clams (steamed) | 3 ounces | 54.4 |
| Halibut (Atlantic and Pacific, cooked, dry heat) | 3 ounces | 47.1 |
| Shrimp (steamed) | 3 ounces | 42.1 |
| Noodles (egg, cooked, enriched) | 1 cup | 38.2 |
| Crab (queen, steamed) | 3 ounces | 37.7 |
| Chicken (light-meat, roasted) | 1 cup | 36.1 |
| Pork (tenderloin, roasted) | 3 ounces | 32.5 |
| Salmon (sockeye, cooked, dry heat) | 3 ounces | 30.2 |
| Beef (plate steak, grilled) | 3 ounces | 28.9 |
| Sunflowers seeds (dried) | ¼ cup | 18.6 |
| Whole-wheat bread | 2 slices | 16.4 |
| Rice (brown, long-grain, cooked) | 1 cup | 11.7 |
| Milk (fat free or skim) | 8 fl oz. (1 cup) | 7.6 |
| *Above the tolerable upper intake level (UL) of 400 μg/day. | ||
Selenium supplements are available in several forms and mostly unregulated by the FDA. Sodium selenite and sodium selenate are inorganic forms of selenium. Sodium selenate is almost completely absorbed, but a significant amount is excreted in the urine before it can be incorporated into proteins. Sodium selenite is only about 50% absorbed but is better retained than selenate once it is absorbed. Selenomethionine, an organic form of selenium, is about 90% absorbed (65); however, only about 34% may then actually be converted to free selenomethionine (139). Selenomethionine and selenium-enriched yeast, which mainly supply selenomethionine, are also available as supplements. The consumer should be aware that some forms of selenium-enriched yeast on the market contain mainly inorganic forms of selenium added to the yeast.
Humans can metabolize both inorganic and organic forms of selenium to selenocysteine and incorporate into selenoenzymes. In intervention trials, supplementation with selenomethionine more effectively increased blood selenium concentrations compared to supplementation with inorganic forms (i.e., sodium selenite and sodium selenate) (138). Yet, inorganic forms may increase plasma glutathione peroxidase (GPx) activity more effectively than organic forms (reviewed in 140). It has also been suggested that the incorporation of selenomethionine in place of methionine into tissue proteins may ensure that selenium is available upon protein turnover (138).
Selenium-enriched foods have been of interest to scientists, especially because of the suggestion that some of the chemical forms of selenium produced by plants might be more potent modifiers of cancer risk than those currently available in supplements. Although there is currently no evidence of long-term health benefits associated with the consumption of selenium-enriched foods, results from animal studies and intervention trials suggest that protein-based sources of selenium are more effective at increasing GPx activity than selenium-enriched yeast and selenomethionine (140). Food fortification may also represent a cost-effective strategy to improve selenium nutritional status in populations at risk of inadequacy (141).
Although selenium is required for health, high doses of selenium — especially with long-term supplementation — can be toxic. Acute and fatal toxicities have occurred with accidental or suicidal ingestion of gram quantities of selenium. Clinically significant selenium toxicity was reported in 13 individuals after taking supplements that contained 27.3 mg (27,300 μg) per tablet due to a manufacturing error. Chronic selenium toxicity (selenosis) may occur with smaller doses of selenium over long periods of time. The most common symptoms of selenosis are hair and nail brittleness and loss (65, 142). Other symptoms may include gastrointestinal disturbances, skin rashes, a garlic breath odor, fatigue, irritability, and neurological disorders. In an area of China with a high prevalence of selenosis, toxic effects occurred with increasing frequency when blood selenium concentrations reached a level corresponding to an intake of 850 μg/day.
The Food and Nutrition Board of the US Institute of Medicine (now the National Academy of Medicine) set the tolerable upper intake level (UL) for selenium at 400 μg/day in adults based on the prevention of hair and nail brittleness and loss and early signs of chronic selenium toxicity (65). The UL includes both selenium obtained from food and selenium from supplements (Table 3).
A few studies have examined the relationship between selenium status and type 2 diabetes mellitus. In the cross-sectional analysis of NHANES III (1988-1994) data from 8,876 adult participants, the highest versus lowest quintile of serum selenium concentrations (≥137 μg/L vs. <111 μg/L) was associated with an increased risk of type 2 diabetes (143). Data analyses from 917 participants (≥40 years of age) of NHANES 2003-2004 also indicated an increased prevalence of type 2 diabetes in the highest versus lowest quartile of serum selenium concentrations (≥147 μg/L vs. <124 μg/L). Individuals in the highest versus lowest quartile of serum selenium concentrations also had higher levels of plasma glucose and glycated hemoglobin (HbA1c), suggestive of poor glycemic control (144). In NHANES 2013-2016, a cross-sectional analysis of 2,706 adults with normal glucose metabolism, the highest (≥209 μg/L) versus lowest (<181 μg/L) quartile of blood selenium concentration were associated with elevated markers of glucose metabolism, including fasting plasma glucose concentration, HbA1c, and circulating insulin (145). In a dose-response meta-analysis of 34 observational studies, circulating selenium concentration of at least 160 μg/L was associated with a near doubling of risk for type 2 diabetes when compared to a circulating concentration of 90 μg/L; a significantly higher risk was also observed at 120 μg/L or greater (RR, 1.27; 95% CI, 1.10-1.47). In this analysis, dietary selenium intakes greater than 80 μg/day were linked to a higher risk of type 2 diabetes compared to dietary selenium intake of 55 μg/day (146).
Type 2 diabetes has been evaluated in randomized controlled trials as a secondary endpoint. The randomized, double-blind, placebo-controlled study in 1,312 participants in the Nutritional Prevention of Cancer (NPC) trial found that selenium supplementation (200 μg/day; mean follow-up of 7.7 years) significantly increased the risk for type 2 diabetes in participants in the highest tertile of baseline plasma selenium concentration (147). In addition, in the Selenium and Vitamin E Cancer Prevention Trial (SELECT), more cases of type 2 diabetes were found in the selenium group (200 μg/day; median follow-up of 5.5 years) than in the placebo group, but this was only a trend and not statistically significant (87). However, other randomized controlled trials have reported 200 μg/day for ~3-4 years does not increase diabetes risk (88, 148).
At present, the mechanisms behind some of these observations are not well understood. An increase in insulin sensitivity has been reported in individuals with congenital (inborn) deficiency of most selenoproteins (149). Results from several animal studies also indicated that selenium supplementation and selenoproteins may interfere with insulin action and glucose homeostasis (reviewed in 150). On the other hand, studies have found that impaired glucose metabolism in patients with type 2 diabetes may affect SELENOP expression and selenium homeostasis (15, 151, 152). While more research is needed to fully understand the interplay between carbohydrate metabolism and selenium homeostasis, the use of high-dose selenium supplements in healthy individuals are currently discouraged in those with high selenium status and/or at increased risk for developing type 2 diabetes (150, 153, 154).
At present, few interactions between selenium and medications have been reported (155). For example, the anticonvulsant drug valproic acid and the chemotherapeutic agent cisplatin may lower circulating selenium concentrations in treated subjects (156, 157). Also, supplemental sodium selenite was found to reduce the toxicity of the antibiotic nitrofurantoin and the herbicide paraquat in animal studies (158).
A three-year randomized controlled trial in 160 patients with documented coronary artery disease (CAD) and low high-density-lipoprotein (HDL) levels found that a combination of simvastatin (Zocor) and niacin increased HDL2 subfraction levels, inhibited the progression of coronary artery stenosis (narrowing), and decreased the frequency of cardiovascular events (159). Surprisingly, when an antioxidant combination (1,000 mg of vitamin C, 800 IU of vitamin E (d-α-tocopherol), 100 μg of selenium, and 25 mg of β-carotene daily) was taken with the simvastatin-niacin combination, the protective effects were diminished. However, the individual contribution of selenium cannot be established, and other studies have reported that antioxidant vitamins alone could interfere with the action of HDL-raising drugs, including statins (160).
The average American diet is estimated to provide about 100 μg/day of selenium, an amount that is well above the current RDA (55 μg/day for adults) and appears sufficient to optimize plasma and cellular glutathione peroxidase (GPx) activity. A 10-week randomized controlled study in healthy British adults (ages, 50-64 years) estimated that about 105 μg/day of total selenium intake was required to maximize the plasma concentrations of selenoprotein P (SELENOP), another useful biomarker of selenium status (161). However, a similar trial in an American cohort with higher baseline plasma selenium concentrations found no effect of selenium supplementation on SELENOP concentrations (162). The amount of selenium in multivitamin/mineral (MVM) supplements varies considerably: most MVMs on the market in the US provide the Daily Value (DV) of 55 μg or a lower amount, but some contain up to 200 μg of selenium (163). Eating a varied diet and taking a daily MVM supplement should provide sufficient selenium for most people in the US and help improve selenium status in populations with lower selenium intakes outside the US.
At present, the effect of selenium supplementation on cancer risk is not clear enough to support a general recommendation for an extra selenium supplement, especially in men with serum selenium concentrations consistent with adequate selenium intakes. SELECT found that 200 μg/day of selenium did not reduce the risk of prostate cancer (87), refuting the results of the NPC trial (see Cancer). Another multicenter, randomized, double-blind, placebo-controlled trial (the Negative Biopsy Trial) in 699 men at high risk of prostate cancer found no effect of either 200 μg/day or 400 μg/day of selenium on prostate cancer risk during a mean follow-up of 36 months (164). In addition, because current evidence suggests a U-shaped relationship between selenium status and prostate cancer risk (165), men with replete selenium status should avoid taking supplemental selenium that would exceed 200 μg/day.
At present, there is no clinical evidence showing that selenium supplementation above recommended levels decreases the risk of breast cancer, although some, but not all, observational studies have found an inverse relationship between selenium status and breast cancer in women (72).
The RDA of selenium for older adults is the same as for younger adults: 55 μg/day. A five-year, randomized, double-blind, placebo-controlled trial in healthy Danish older people (ages at inclusion, 60-74 years) found that selenium supplementation (100-300 μg/day) had little-to-no impact on circulating levels of antioxidant enzymes, including GPx (166).
Originally written in 2001 by:
Jane Higdon, Ph.D.
Linus Pauling Institute
Oregon State University
Updated in October 2003 by:
Jane Higdon, Ph.D.
Linus Pauling Institute
Oregon State University
Updated in November 2007 by:
Victoria J. Drake, Ph.D.
Linus Pauling Institute
Oregon State University
Updated in November 2014 by:
Barbara Delage, Ph.D.
Linus Pauling Institute
Oregon State University
Updated in November 2024 by:
Victoria J. Drake, Ph.D.
Linus Pauling Institute
Oregon State University
Reviewed in February 2024 by:
Petra A. Tsuji, Ph.D., M.P.H.
Professor
Department of Biological Sciences
Towson University
Copyright 2001-2024 Linus Pauling Institute
1. Rayman MP. The importance of selenium to human health. Lancet. 2000;356(9225):233-241. (PubMed)
2. Mangiapane E, Pessione A, Pessione E. Selenium and selenoproteins: an overview on different biological systems. Curr Protein Pept Sci. 2014;15(6):598-607. (PubMed)
3. Burk RF, Hill KE. Regulation of selenium metabolism and transport. Annu Rev Nutr. 2015;35:109-134. (PubMed)
4. Berry MJ, Banu L, Harney JW, Larsen PR. Functional characterization of the eukaryotic SECIS elements which direct selenocysteine insertion at UGA codons. EMBO J. 1993;12(8):3315-3322. (PubMed)
5. Mariotti M, Ridge PG, Zhang Y, et al. Composition and evolution of the vertebrate and mammalian selenoproteomes. PLoS One. 2012;7(3):e33066. (PubMed)
6. Terry EN, Diamond JA. Selenium. In: Erdman Jr J, Macdonald J, Zeisel S, eds. Present Knowledge in Nutrition: John Wiley & Sons, Inc; 2012:568-585.
7. Boitani C, Puglisi R. Selenium, a key element in spermatogenesis and male fertility. Adv Exp Med Biol. 2008;636:65-73. (PubMed)
8. Arner ES. Focus on mammalian thioredoxin reductases--important selenoproteins with versatile functions. Biochim Biophys Acta. 2009;1790(6):495-526. (PubMed)
9. Lu J, Holmgren A. The thioredoxin antioxidant system. Free Radic Biol Med. 2014;66:75-87. (PubMed)
10. Schomburg L. Selenium, selenoproteins and the thyroid gland: interactions in health and disease. Nat Rev Endocrinol. 2012;8(3):160-171. (PubMed)
11. Tsuji PA, Santesmasses D, Lee BJ, Gladyshev VN, Hatfield DL. Historical roles of selenium and selenoproteins in health and development: the good, the bad and the ugly. Int J Mol Sci. 2021;23(1):5. (PubMed)
12. Hill KE, Wu S, Motley AK, et al. Production of selenoprotein P (Sepp1) by hepatocytes is central to selenium homeostasis. J Biol Chem. 2012;287(48):40414-40424. (PubMed)
13. Schomburg L. Selenoprotein P - Selenium transport protein, enzyme and biomarker of selenium status. Free Radic Biol Med. 2022;191:150-163. (PubMed)
14. Olson GE, Winfrey VP, Hill KE, Burk RF. Megalin mediates selenoprotein P uptake by kidney proximal tubule epithelial cells. J Biol Chem. 2008;283(11):6854-6860. (PubMed)
15. Misu H, Takamura T, Takayama H, et al. A liver-derived secretory protein, selenoprotein P, causes insulin resistance. Cell Metab. 2010;12(5):483-495. (PubMed)
16. Jujic A, Molvin J, Schomburg L, et al. Selenoprotein P deficiency is associated with higher risk of incident heart failure. Free Radic Biol Med. 2023;207:11-16. (PubMed)
17. Whanger PD. Selenoprotein expression and function-selenoprotein W. Biochim Biophys Acta. 2009;1790(11):1448-1452. (PubMed)
18. Jeong D, Kim TS, Chung YW, Lee BJ, Kim IY. Selenoprotein W is a glutathione-dependent antioxidant in vivo. FEBS Lett. 2002;517(1-3):225-228. (PubMed)
19. Reeves MA, Hoffmann PR. The human selenoproteome: recent insights into functions and regulation. Cell Mol Life Sci. 2009;66(15):2457-2478. (PubMed)
20. Reszka E, Jablonska E, Gromadzinska J, Wasowicz W. Relevance of selenoprotein transcripts for selenium status in humans. Genes Nutr. 2012;7(2):127-137. (PubMed)
21. Chung YW, Jeong D, Noh OJ, et al. Antioxidative role of selenoprotein W in oxidant-induced mouse embryonic neuronal cell death. Mol Cells. 2009;27(5):609-613. (PubMed)
22. Jeon YH, Park YH, Kwon JH, Lee JH, Kim IY. Inhibition of 14-3-3 binding to Rictor of mTORC2 for Akt phosphorylation at Ser473 is regulated by selenoprotein W. Biochim Biophys Acta. 2013;1833(10):2135-2142. (PubMed)
23. Kanai F, Marignani PA, Sarbassova D, et al. TAZ: a novel transcriptional co-activator regulated by interactions with 14-3-3 and PDZ domain proteins. EMBO J. 2000;19(24):6778-6791. (PubMed)
24. Jeon YH, Park YH, Lee JH, Hong JH, Kim IY. Selenoprotein W enhances skeletal muscle differentiation by inhibiting TAZ binding to 14-3-3 protein. Biochim Biophys Acta. 2014;1843(7):1356-1364. (PubMed)
25. Alkan Z, Duong FL, Hawkes WC. Selenoprotein W controls epidermal growth factor receptor surface expression, activation and degradation via receptor ubiquitination. Biochim Biophys Acta. 2015;1853(5):1087-1095. (PubMed)
26. Ganichkin OM, Xu XM, Carlson BA, et al. Structure and catalytic mechanism of eukaryotic selenocysteine synthase. J Biol Chem. 2008;283(9):5849-5865. (PubMed)
27. Lee BC, Peterfi Z, Hoffmann FW, et al. MsrB1 and MICALs regulate actin assembly and macrophage function via reversible stereoselective methionine oxidation. Mol Cell. 2013;51(3):397-404. (PubMed)
28. Kim HY. The methionine sulfoxide reduction system: selenium utilization and methionine sulfoxide reductase enzymes and their functions. Antioxid Redox Signal. 2013;19(9):958-969. (PubMed)
29. Kumaraswamy E, Malykh A, Korotkov KV, et al. Structure-expression relationships of the 15-kDa selenoprotein gene. Possible role of the protein in cancer etiology. J Biol Chem. 2000;275(45):35540-35547. (PubMed)
30. Korotkov KV, Kumaraswamy E, Zhou Y, Hatfield DL, Gladyshev VN. Association between the 15-kDa selenoprotein and UDP-glucose:glycoprotein glucosyltransferase in the endoplasmic reticulum of mammalian cells. J Biol Chem. 2001;276(18):15330-15336. (PubMed)
31. Labunskyy VM, Ferguson AD, Fomenko DE, Chelliah Y, Hatfield DL, Gladyshev VN. A novel cysteine-rich domain of Sep15 mediates the interaction with UDP-glucose:glycoprotein glucosyltransferase. J Biol Chem. 2005;280(45):37839-37845. (PubMed)
32. Labunskyy VM, Hatfield DL, Gladyshev VN. The Sep15 protein family: roles in disulfide bond formation and quality control in the endoplasmic reticulum. IUBMB Life. 2007;59(1):1-5. (PubMed)
33. Kasaikina MV, Fomenko DE, Labunskyy VM, et al. Roles of the 15-kDa selenoprotein (Sep15) in redox homeostasis and cataract development revealed by the analysis of Sep 15 knockout mice. J Biol Chem. 2011;286(38):33203-33212. (PubMed)
34. Hatfield DL, Tsuji PA, Carlson BA, Gladyshev VN. Selenium and selenocysteine: roles in cancer, health, and development. Trends Biochem Sci. 2014;39(3):112-120. (PubMed)
35. Canter JA, Ernst SE, Peters KM, et al. Selenium and the 15kDa selenoprotein impact colorectal tumorigenesis by modulating intestinal barrier integrity. Int J Mol Sci. 2021;22(19):10651. (PubMed)
36. Curran JE, Jowett JB, Elliott KS, et al. Genetic variation in selenoprotein S influences inflammatory response. Nat Genet. 2005;37(11):1234-1241. (PubMed)
37. Santos LR, Duraes C, Mendes A, et al. A polymorphism in the promoter region of the selenoprotein S gene (SEPS1) contributes to Hashimoto's thyroiditis susceptibility. J Clin Endocrinol Metab. 2014;99(4):E719-723. (PubMed)
38. Alanne M, Kristiansson K, Auro K, et al. Variation in the selenoprotein S gene locus is associated with coronary heart disease and ischemic stroke in two independent Finnish cohorts. Hum Genet. 2007;122(3-4):355-365. (PubMed)
39. Cox AJ, Lehtinen AB, Xu J, et al. Polymorphisms in the selenoprotein S gene and subclinical cardiovascular disease in the Diabetes Heart Study. Acta Diabetol. 2013;50(3):391-399. (PubMed)
40. Moses EK, Johnson MP, Tommerdal L, et al. Genetic association of preeclampsia to the inflammatory response gene SEPS1. Am J Obstet Gynecol. 2008;198(3):336 e331-335. (PubMed)
41. Shibata T, Arisawa T, Tahara T, et al. Selenoprotein S (SEPS1) gene -105G>A promoter polymorphism influences the susceptibility to gastric cancer in the Japanese population. BMC Gastroenterol. 2009;9:2. (PubMed)
42. Shchedrina VA, Zhang Y, Labunskyy VM, Hatfield DL, Gladyshev VN. Structure-function relations, physiological roles, and evolution of mammalian ER-resident selenoproteins. Antioxid Redox Signal. 2010;12(7):839-849. (PubMed)
43. Li X, Hill KE, Burk RF, May JM. Selenium spares ascorbate and alpha-tocopherol in cultured liver cell lines under oxidant stress. FEBS Lett. 2001;508(3):489-492. (PubMed)
44. May JM, Mendiratta S, Hill KE, Burk RF. Reduction of dehydroascorbate to ascorbate by the selenoenzyme thioredoxin reductase. J Biol Chem. 1997;272(36):22607-22610. (PubMed)
45. Murer SB, Aeberli I, Braegger CP, et al. Antioxidant supplements reduced oxidative stress and stabilized liver function tests but did not reduce inflammation in a randomized controlled trial in obese children and adolescents. J Nutr. 2014;144(2):193-201. (PubMed)
46. Schneider MJ, Fiering SN, Thai B, et al. Targeted disruption of the type 1 selenodeiodinase gene (Dio1) results in marked changes in thyroid hormone economy in mice. Endocrinology. 2006;147(1):580-589. (PubMed)
47. Hess SY. The impact of common micronutrient deficiencies on iodine and thyroid metabolism: the evidence from human studies. Best Pract Res Clin Endocrinol Metab. 2010;24(1):117-132. (PubMed)
48. Thomson CD. Assessment of requirements for selenium and adequacy of selenium status: a review. Eur J Clin Nutr. 2004;58(3):391-402. (PubMed)
49. Shahmiri SS, Eghbali F, Ismaeil A, et al. Selenium deficiency after bariatric surgery, incidence and symptoms: a systematic review and meta-analysis. Obes Surg. 2022;32(5):1719-1725. (PubMed)
50. Yan W, Meihao W, Zihan S, et al. Correlation between Crohn's disease activity and serum selenium concentration. Clin Ther. 2022;44(5):736-743 e3. (PubMed)
51. Cooper A, Mones RL, Heird WC. Nutritional management of infants and children with specific diseases and other conditions. In: Ross AC, Caballero B, Cousins RJ, Tucker KL, Ziegler TR, eds. Modern Nutrition in Health and Disease. 11th ed. Baltimore: Lippincott Williams & Wilkins; 2014:988-1005.
52. Sunde RA. Selenium. In: Ross AC, Caballero B, Cousins RJ, Tucker KL, Ziegler TR, eds. Modern Nutrition in Health and Disease. 11th ed. Baltimore: Lippincott Williams & Wilkins; 2014:265-276.
53. Lei C, Niu X, Wei J, Zhu J, Zhu Y. Interaction of glutathione peroxidase-1 and selenium in endemic dilated cardiomyopathy. Clin Chim Acta. 2009;399(1-2):102-108. (PubMed)
54. Zhou H, Wang T, Li Q, Li D. Prevention of Keshan disease by selenium supplementation: a systematic review and meta-analysis. Biol Trace Elem Res. 2018;186(1):98-105. (PubMed)
55. Chen J. An original discovery: selenium deficiency and Keshan disease (an endemic heart disease). Asia Pac J Clin Nutr. 2012;21(3):320-326. (PubMed)
56. Beck MA, Kolbeck PC, Rohr LH, Shi Q, Morris VC, Levander OA. Benign human enterovirus becomes virulent in selenium-deficient mice. J Med Virol. 1994;43(2):166-170. (PubMed)
57. Harthill M. Review: micronutrient selenium deficiency influences evolution of some viral infectious diseases. Biol Trace Elem Res. 2011;143(3):1325-1336. (PubMed)
58. Stone R. Diseases. A medical mystery in middle China. Science. 2009;324(5933):1378-1381. (PubMed)
59. Wang K, Yu J, Liu H, et al. Endemic Kashin-Beck disease: A food-sourced osteoarthropathy. Semin Arthritis Rheum. 2020;50(2):366-372. (PubMed)
60. Du XH, Dai XX, Xia Song R, et al. SNP and mRNA expression for glutathione peroxidase 4 in Kashin-Beck disease. Br J Nutr. 2012;107(2):164-169. (PubMed)
61. Xiong YM, Mo XY, Zou XZ, et al. Association study between polymorphisms in selenoprotein genes and susceptibility to Kashin-Beck disease. Osteoarthritis Cartilage. 2010;18(6):817-824. (PubMed)
62. Yu FF, Sun L, Zhou GY, Ping ZG, Guo X, Ba Y. Meta-analysis of association studies of selenoprotein gene polymorphism and Kashin-Beck disease: an updated systematic review. Biol Trace Elem Res. 2022;200(2):543-550. (PubMed)
63. Zou K, Liu G, Wu T, Du L. Selenium for preventing Kashin-Beck osteoarthropathy in children: a meta-analysis. Osteoarthritis Cartilage. 2009;17(2):144-151. (PubMed)
64. Jirong Y, Huiyun P, Zhongzhe Y, et al. Sodium selenite for treatment of Kashin-Beck disease in children: a systematic review of randomised controlled trials. Osteoarthritis Cartilage. 2012;20(7):605-613. (PubMed)
65. Food and Nutrition Board, Institute of Medicine. Selenium. Dietary reference intakes for vitamin C, vitamin E, selenium, and carotenoids. Washington, D.C.: National Academy Press; 2000:284-324. (National Academy Press)
66. Combs GF, Jr., Gray WP. Chemopreventive agents: selenium. Pharmacol Ther. 1998;79(3):179-192. (PubMed)
67. Vinceti M, Dennert G, Crespi CM, et al. Selenium for preventing cancer. Cochrane Database Syst Rev. 2014;3:CD005195. (PubMed)
68. Hansen RD, Albieri V, Tjonneland A, Overvad K, Andersen KK, Raaschou-Nielsen O. Effects of smoking and antioxidant micronutrients on risk of colorectal cancer. Clin Gastroenterol Hepatol. 2013;11(4):406-15.e3. (PubMed)
69. Northrop-Clewes CA, Thurnham DI. Monitoring micronutrients in cigarette smokers. Clin Chim Acta. 2007;377(1-2):14-38. (PubMed)
70. Hazane-Puch F, Champelovier P, Arnaud J, et al. Long-term selenium supplementation in HaCaT cells: importance of chemical form for antagonist (protective versus toxic) activities. Biol Trace Elem Res. 2013;154(2):288-298. (PubMed)
71. Weekley CM, Harris HH. Which form is that? The importance of selenium speciation and metabolism in the prevention and treatment of disease. Chem Soc Rev. 2013;42(23):8870-8894. (PubMed)
72. Babaknejad N, Sayehmiri F, Sayehmiri K, et al. The relationship between selenium levels and breast cancer: a systematic review and meta-analysis. Biol Trace Elem Res. 2014;159(1-3):1-7. (PubMed)
73. Hao R, Yu P, Gui L, Wang N, Pan D, Wang S. Relationship between serum levels of selenium and thyroid cancer: a systematic review and meta-analysis. Nutr Cancer. 2023;75(1):14-23. (PubMed)
74. Meplan C, Hesketh J. The influence of selenium and selenoprotein gene variants on colorectal cancer risk. Mutagenesis. 2012;27(2):177-186. (PubMed)
75. Li J, Zhu Y, Zhou Y, et al. The SELS rs34713741 polymorphism is associated with susceptibility to colorectal cancer and gastric cancer: a meta-analysis. Genet Test Mol Biomarkers. 2020;24(12):835-844. (PubMed)
76. Steinbrecher A, Meplan C, Hesketh J, et al. Effects of selenium status and polymorphisms in selenoprotein genes on prostate cancer risk in a prospective study of European men. Cancer Epidemiol Biomarkers Prev. 2010;19(11):2958-2968. (PubMed)
77. Gerstenberger JP, Bauer SR, Van Blarigan EL, et al. Selenoprotein and antioxidant genes and the risk of high-grade prostate cancer and prostate cancer recurrence. Prostate. 2015;75(1):60-69. (PubMed)
78. Meplan C, Rohrmann S, Steinbrecher A, et al. Polymorphisms in thioredoxin reductase and selenoprotein K genes and selenium status modulate risk of prostate cancer. PLoS One. 2012;7(11):e48709. (PubMed)
79. Penney KL, Schumacher FR, Li H, et al. A large prospective study of SEP15 genetic variation, interaction with plasma selenium levels, and prostate cancer risk and survival. Cancer Prev Res (Phila). 2010;3(5):604-610. (PubMed)
80. Yu SY, Zhu YJ, Li WG. Protective role of selenium against hepatitis B virus and primary liver cancer in Qidong. Biol Trace Elem Res. 1997;56(1):117-124. (PubMed)
81. Duffield-Lillico AJ, Dalkin BL, Reid ME, et al. Selenium supplementation, baseline plasma selenium status and incidence of prostate cancer: an analysis of the complete treatment period of the Nutritional Prevention of Cancer Trial. BJU Int. 2003;91(7):608-612. (PubMed)
82. Clark LC, Combs GF, Jr., Turnbull BW, et al. Effects of selenium supplementation for cancer prevention in patients with carcinoma of the skin. A randomized controlled trial. Nutritional Prevention of Cancer Study Group. JAMA. 1996;276(24):1957-1963. (PubMed)
83. Reid ME, Duffield-Lillico AJ, Slate E, et al. The nutritional prevention of cancer: 400 mcg per day selenium treatment. Nutr Cancer. 2008;60(2):155-163. (PubMed)
84. National Cancer Institute. Review of Prostate Cancer Prevention Study Shows No Benefit for Use of Selenium and Vitamin E Supplements. [Web page]. http://www.cancer.gov/newscenter/pressreleases/SELECTresults2008. Accessed 10/28/08.
85. Hatfield DL, Gladyshev VN. The Outcome of Selenium and Vitamin E Cancer Prevention Trial (SELECT) reveals the need for better understanding of selenium biology. Mol Interv. 2009;9(1):18-21. (PubMed)
86. Klein EA, Thompson IM, Jr., Tangen CM, et al. Vitamin E and the risk of prostate cancer: the Selenium and Vitamin E Cancer Prevention Trial (SELECT). JAMA. 2011;306(14):1549-1556. (PubMed)
87. Lippman SM, Klein EA, Goodman PJ, et al. Effect of selenium and vitamin E on risk of prostate cancer and other cancers: the Selenium and Vitamin E Cancer Prevention Trial (SELECT). JAMA. 2009;301(1):39-51. (PubMed)
88. Thompson PA, Ashbeck EL, Roe DJ, et al. Selenium supplementation for prevention of colorectal adenomas and risk of associated type 2 diabetes. J Natl Cancer Inst. 2016;108(12):djw152. (PubMed)
89. Vinceti M, Crespi CM, Malagoli C, Del Giovane C, Krogh V. Friend or foe? The current epidemiologic evidence on selenium and human cancer risk. J Environ Sci Health C Environ Carcinog Ecotoxicol Rev. 2013;31(4):305-341. (PubMed)
90. Flores-Mateo G, Carrillo-Santisteve P, Elosua R, et al. Antioxidant enzyme activity and coronary heart disease: meta-analyses of observational studies. Am J Epidemiol. 2009;170(2):135-147. (PubMed)
91. Bleys J, Navas-Acien A, Guallar E. Serum selenium levels and all-cause, cancer, and cardiovascular mortality among US adults. Arch Intern Med. 2008;168(4):404-410. (PubMed)
92. Eaton CB, Abdul Baki AR, Waring ME, Roberts MB, Lu B. The association of low selenium and renal insufficiency with coronary heart disease and all-cause mortality: NHANES III follow-up study. Atherosclerosis. 2010;212(2):689-694. (PubMed)
93. Shi W, Su L, Wang J, Wang F, Liu X, Dou J. Correlation between dietary selenium intake and stroke in the National Health and Nutrition Examination Survey 2003-2018. Ann Med. 2022;54(1):1395-1402. (PubMed)
94. Ding J, Zhang Y. Relationship between the circulating selenium level and stroke: a meta-analysis of observational studies. J Am Nutr Assoc. 2022;41(5):444-452. (PubMed)
95. Laclaustra M, Navas-Acien A, Stranges S, Ordovas JM, Guallar E. Serum selenium concentrations and hypertension in the US Population. Circ Cardiovasc Qual Outcomes. 2009;2(4):369-376. (PubMed)
96. Kuruppu D, Hendrie HC, Yang L, Gao S. Selenium levels and hypertension: a systematic review of the literature. Public Health Nutr. 2014;17(6):1342-1352. (PubMed)
97. Laclaustra M, Stranges S, Navas-Acien A, Ordovas JM, Guallar E. Serum selenium and serum lipids in US adults: National Health and Nutrition Examination Survey (NHANES) 2003-2004. Atherosclerosis. 2010;210(2):643-648. (PubMed)
98. Stranges S, Laclaustra M, Ji C, et al. Higher selenium status is associated with adverse blood lipid profile in British adults. J Nutr. 2010;140(1):81-87. (PubMed)
99. Rees K, Hartley L, Day C, Flowers N, Clarke A, Stranges S. Selenium supplementation for the primary prevention of cardiovascular disease. Cochrane Database Syst Rev. 2013;1:CD009671. (PubMed)
100. Flores-Mateo G, Navas-Acien A, Pastor-Barriuso R, Guallar E. Selenium and coronary heart disease: a meta-analysis. Am J Clin Nutr. 2006;84(4):762-773. (PubMed)
101. Jenkins DJA, Kitts D, Giovannucci EL, et al. Selenium, antioxidants, cardiovascular disease, and all-cause mortality: a systematic review and meta-analysis of randomized controlled trials. Am J Clin Nutr. 2020;112(6):1642-1652. (PubMed)
102. Kuria A, Tian H, Li M, et al. Selenium status in the body and cardiovascular disease: a systematic review and meta-analysis. Crit Rev Food Sci Nutr. 2021;61(21):3616-3625. (PubMed)
103. McKenzie RC, Beckett GJ, Arthur JR. Effects of selenium on immunity and aging. In: Hatfield DL, Berry MJ, Gladyshev VN, eds. Selenium: Its Molecular Biology and Role in Human Health. 2nd ed. New York: Springer; 2006:311-323.
104. Huang Z, Rose AH, Hoffmann PR. The role of selenium in inflammation and immunity: from molecular mechanisms to therapeutic opportunities. Antioxid Redox Signal. 2012;16(7):705-743. (PubMed)
105. Fairweather-Tait SJ, Filippini T, Vinceti M. Selenium status and immunity. Proc Nutr Soc. 2023;82(1):32-38. (PubMed)
106. Mattmiller SA, Carlson BA, Sordillo LM. Regulation of inflammation by selenium and selenoproteins: impact on eicosanoid biosynthesis. J Nutr Sci. 2013;2:e28. (PubMed)
107. Norton RL, Hoffmann PR. Selenium and asthma. Mol Aspects Med. 2012;33(1):98-106. (PubMed)
108. Speckmann B, Steinbrenner H. Selenium and selenoproteins in inflammatory bowel diseases and experimental colitis. Inflamm Bowel Dis. 2014;20(6):1110-1119. (PubMed)
109. Drain PK, Kupka R, Mugusi F, Fawzi WW. Micronutrients in HIV-positive persons receiving highly active antiretroviral therapy. Am J Clin Nutr. 2007;85(2):333-345. (PubMed)
110. Stone CA, Kawai K, Kupka R, Fawzi WW. Role of selenium in HIV infection. Nutr Rev. 2010;68(11):671-681. (PubMed)
111. de Menezes Barbosa EG, Junior FB, Machado AA, Navarro AM. A longer time of exposure to antiretroviral therapy improves selenium levels. Clin Nutr. 2015;34(2):248-51. (PubMed)
112. Baum MK, Miguez-Burbano MJ, Campa A, Shor-Posner G. Selenium and interleukins in persons infected with human immunodeficiency virus type 1. J Infect Dis. 2000;182 Suppl 1:S69-73. (PubMed)
113. Kalantari P, Narayan V, Natarajan SK, et al. Thioredoxin reductase-1 negatively regulates HIV-1 transactivating protein Tat-dependent transcription in human macrophages. J Biol Chem. 2008;283(48):33183-33190. (PubMed)
114. Burbano X, Miguez-Burbano MJ, McCollister K, et al. Impact of a selenium chemoprevention clinical trial on hospital admissions of HIV-infected participants. HIV Clin Trials. 2002;3(6):483-491. (PubMed)
115. Hurwitz BE, Klaus JR, Llabre MM, et al. Suppression of human immunodeficiency virus type 1 viral load with selenium supplementation: a randomized controlled trial. Arch Intern Med. 2007;167(2):148-154. (PubMed)
116. Kupka R, Mugusi F, Aboud S, Hertzmark E, Spiegelman D, Fawzi WW. Effect of selenium supplements on hemoglobin concentration and morbidity among HIV-1-infected Tanzanian women. Clin Infect Dis. 2009;48(10):1475-1478. (PubMed)
117. Kupka R, Mugusi F, Aboud S, et al. Randomized, double-blind, placebo-controlled trial of selenium supplements among HIV-infected pregnant women in Tanzania: effects on maternal and child outcomes. Am J Clin Nutr. 2008;87(6):1802-1808. (PubMed)
118. Baum MK, Campa A, Lai S, et al. Effect of micronutrient supplementation on disease progression in asymptomatic, antiretroviral-naive, HIV-infected adults in Botswana: a randomized clinical trial. JAMA. 2013;310(20):2154-2163. (PubMed)
119. Muzembo BA, Ngatu NR, Januka K, et al. Selenium supplementation in HIV-infected individuals: A systematic review of randomized controlled trials. Clin Nutr ESPEN. 2019;34:1-7. (PubMed)
120. Bone RC, Balk RA, Cerra FB, et al. Definitions for sepsis and organ failure and guidelines for the use of innovative therapies in sepsis. The ACCP/SCCM Consensus Conference Committee. American College of Chest Physicians/Society of Critical Care Medicine. Chest. 1992;101(6):1644-1655. (PubMed)
121. Mann EA, Baun MM, Meininger JC, Wade CE. Comparison of mortality associated with sepsis in the burn, trauma, and general intensive care unit patient: a systematic review of the literature. Shock. 2012;37(1):4-16. (PubMed)
122. Alhazzani W, Jacobi J, Sindi A, et al. The effect of selenium therapy on mortality in patients with sepsis syndrome: a systematic review and meta-analysis of randomized controlled trials. Crit Care Med. 2013;41(6):1555-1564. (PubMed)
123. Huang TS, Shyu YC, Chen HY, et al. Effect of parenteral selenium supplementation in critically ill patients: a systematic review and meta-analysis. PLoS One. 2013;8(1):e54431. (PubMed)
124. Bloos F, Trips E, Nierhaus A, et al. Effect of sodium selenite administration and procalcitonin-guided therapy on mortality in patients with severe sepsis or septic shock: a randomized clinical trial. JAMA Intern Med. 2016;176(9):1266-1276. (PubMed)
125. Rhodes A, Evans LE, Alhazzani W, et al. Surviving Sepsis Campaign: International guidelines for management of sepsis and septic shock: 2016. Crit Care Med. 2017;45(3):486-552. (PubMed)
126. Xu S, Huang H, Qian J, et al. Prevalence of Hashimoto thyroiditis in adults with papillary thyroid cancer and its association with cancer recurrence and outcomes. JAMA Netw Open. 2021;4(7):e2118526. (PubMed)
127. van Zuuren EJ, Albusta AY, Fedorowicz Z, Carter B, Pijl H. Selenium supplementation for Hashimoto's thyroiditis: summary of a Cochrane systematic review. Eur Thyroid J. 2014;3(1):25-31. (PubMed)
128. Karanikas G, Schuetz M, Kontur S, et al. No immunological benefit of selenium in consecutive patients with autoimmune thyroiditis. Thyroid. 2008;18(1):7-12. (PubMed)
129. Krysiak R, Okopien B. The effect of levothyroxine and selenomethionine on lymphocyte and monocyte cytokine release in women with Hashimoto's thyroiditis. J Clin Endocrinol Metab. 2011;96(7):2206-2215. (PubMed)
130. Negro R, Greco G, Mangieri T, Pezzarossa A, Dazzi D, Hassan H. The influence of selenium supplementation on postpartum thyroid status in pregnant women with thyroid peroxidase autoantibodies. J Clin Endocrinol Metab. 2007;92(4):1263-1268. (PubMed)
131. Turker O, Kumanlioglu K, Karapolat I, Dogan I. Selenium treatment in autoimmune thyroiditis: 9-month follow-up with variable doses. J Endocrinol. 2006;190(1):151-156. (PubMed)
132. Hu Y, Feng W, Chen H, et al. Effect of selenium on thyroid autoimmunity and regulatory T cells in patients with Hashimoto's thyroiditis: A prospective randomized-controlled trial. Clin Transl Sci. 2021;14(4):1390-1402. (PubMed)
133. Marcocci C, Kahaly GJ, Krassas GE, et al. Selenium and the course of mild Graves' orbitopathy. N Engl J Med. 2011;364(20):1920-1931. (PubMed)
134. Watt T, Cramon P, Bjorner JB, et al. Selenium supplementation for patients with Graves' hyperthyroidism (the GRASS trial): study protocol for a randomized controlled trial. Trials. 2013;14:119. (PubMed)
135. Winther KH, Watt T, Bjorner JB, et al. The chronic autoimmune thyroiditis quality of life selenium trial (CATALYST): study protocol for a randomized controlled trial. Trials. 2014;15:115. (PubMed)
136. Peplow D, Edmonds R. Health risks associated with contamination of groundwater by abandoned mines near Twisp in Okanogan County, Washington, USA. Environ Geochem Health. 2004;26(1):69-79. (PubMed)
137. Chang JC, Gutenmann WH, Reid CM, Lisk DJ. Selenium content of Brazil nuts from two geographic locations in Brazil. Chemosphere. 1995;30(4):801-802. (PubMed)
138. Rayman MP, Infante HG, Sargent M. Food-chain selenium and human health: spotlight on speciation. Br J Nutr. 2008;100(2):238-253. (PubMed)
139. Reyes LH, Encinar JR, Marchante-Gayon JM, Alonso JI, Sanz-Medel A. Selenium bioaccessibility assessment in selenized yeast after "in vitro" gastrointestinal digestion using two-dimensional chromatography and mass spectrometry. J Chromatogr A. 2006;1110(1-2):108-116. (PubMed)
140. Bermingham EN, Hesketh JE, Sinclair BR, Koolaard JP, Roy NC. Selenium-enriched foods are more effective at increasing glutathione peroxidase (GPx) activity compared with selenomethionine: a meta-analysis. Nutrients. 2014;6(10):4002-4031. (PubMed)
141. Giacosa A, Faliva MA, Perna S, Minoia C, Ronchi A, Rondanelli M. Selenium fortification of an Italian rice cultivar via foliar fertilization with sodium selenate and its effects on human serum selenium levels and on erythrocyte glutathione peroxidase activity. Nutrients. 2014;6(3):1251-1261. (PubMed)
142. Lei XG, Combs GF, Jr., Sunde RA, Caton JS, Arthington JD, Vatamaniuk MZ. Dietary selenium across species. Annu Rev Nutr. 2022;42:337-375. (PubMed)
143. Bleys J, Navas-Acien A, Guallar E. Serum selenium and diabetes in U.S. adults. Diabetes Care. 2007;30(4):829-834. (PubMed)
144. Laclaustra M, Navas-Acien A, Stranges S, Ordovas JM, Guallar E. Serum selenium concentrations and diabetes in U.S. adults: National Health and Nutrition Examination Survey (NHANES) 2003-2004. Environ Health Perspect. 2009;117(9):1409-1413. (PubMed)
145. Yang J, Chen E, Choi C, et al. Cross-sectional association of blood selenium with glycemic biomarkers among U.S. adults with normoglycemia in the National Health and Nutrition Examination Survey 2013-2016. Nutrients. 2022;14(19):3972. (PubMed)
146. Vinceti M, Filippini T, Wise LA, Rothman KJ. A systematic review and dose-response meta-analysis of exposure to environmental selenium and the risk of type 2 diabetes in nonexperimental studies. Environ Res. 2021;197:111210. (PubMed)
147. Stranges S, Marshall JR, Natarajan R, et al. Effects of long-term selenium supplementation on the incidence of type 2 diabetes: a randomized trial. Ann Intern Med. 2007;147(4):217-223. (PubMed)
148. Karp DD, Lee SJ, Keller SM, et al. Randomized, double-blind, placebo-controlled, phase III chemoprevention trial of selenium supplementation in patients with resected stage I non-small-cell lung cancer: ECOG 5597. J Clin Oncol. 2013;31(33):4179-4187. (PubMed)
149. Schoenmakers E, Agostini M, Mitchell C, et al. Mutations in the selenocysteine insertion sequence-binding protein 2 gene lead to a multisystem selenoprotein deficiency disorder in humans. J Clin Invest. 2010;120(12):4220-4235. (PubMed)
150. Steinbrenner H. Interference of selenium and selenoproteins with the insulin-regulated carbohydrate and lipid metabolism. Free Radic Biol Med. 2013;65:1538-1547. (PubMed)
151. Kaur P, Rizk NM, Ibrahim S, et al. iTRAQ-based quantitative protein expression profiling and MRM verification of markers in type 2 diabetes. J Proteome Res. 2012;11(11):5527-5539. (PubMed)
152. Yang SJ, Hwang SY, Choi HY, et al. Serum selenoprotein P levels in patients with type 2 diabetes and prediabetes: implications for insulin resistance, inflammation, and atherosclerosis. J Clin Endocrinol Metab. 2011;96(8):E1325-1329. (PubMed)
153. Steinbrenner H, Duntas LH, Rayman MP. The role of selenium in type-2 diabetes mellitus and its metabolic comorbidities. Redox Biol. 2022;50:102236. (PubMed)
154. Cardoso BR, Braat S, Graham RM. Selenium status is associated with insulin resistance markers in adults: findings from the 2013 to 2018 National Health and Nutrition Examination Survey (NHANES). Front Nutr. 2021;8:696024. (PubMed)
155. Azrak RG, Cao S, Pendyala L, et al. Efficacy of increasing the therapeutic index of irinotecan, plasma and tissue selenium concentrations is methylselenocysteine dose dependent. Biochem Pharmacol. 2007;73(9):1280-1287. (PubMed)
156. Graf WD, Oleinik OE, Glauser TA, Maertens P, Eder DN, Pippenger CE. Altered antioxidant enzyme activities in children with a serious adverse experience related to valproic acid therapy. Neuropediatrics. 1998;29(4):195-201. (PubMed)
157. Vernie LN, de Goeij JJ, Zegers C, de Vries M, Baldew GS, McVie JG. Cisplatin-induced changes of selenium levels and glutathione peroxidase activities in blood of testis tumor patients. Cancer Lett. 1988;40(1):83-91. (PubMed)
158. Flodin NW. Micronutrient supplements: toxicity and drug interactions. Prog Food Nutr Sci. 1990;14(4):277-331. (PubMed)
159. Brown BG, Zhao XQ, Chait A, et al. Simvastatin and niacin, antioxidant vitamins, or the combination for the prevention of coronary disease. N Engl J Med. 2001;345(22):1583-1592. (PubMed)
160. Tousoulis D, Antoniades C, Stefanadis C. Statins and antioxidant vitamins: should co-administration be avoided? J Am Coll Cardiol. 2006;47(6):1237; author reply 1237-1238. (PubMed)
161. Hurst R, Armah CN, Dainty JR, et al. Establishing optimal selenium status: results of a randomized, double-blind, placebo-controlled trial. Am J Clin Nutr. 2010;91(4):923-931. (PubMed)
162. Burk RF, Norsworthy BK, Hill KE, Motley AK, Byrne DW. Effects of chemical form of selenium on plasma biomarkers in a high-dose human supplementation trial. Cancer Epidemiol Biomarkers Prev. 2006;15(4):804-810. (PubMed)
163. US Department of Health and Human Services, National Institutes of Health, Office of Dietary Supplements. Dietary Supplement Label Database (DSLD). [Internet]. [cited 10/25/2023]. Available from: https://dsld.od.nih.gov.
164. Algotar AM, Stratton MS, Ahmann FR, et al. Phase 3 clinical trial investigating the effect of selenium supplementation in men at high-risk for prostate cancer. Prostate. 2013;73(3):328-335. (PubMed)
165. Chiang EC, Shen S, Kengeri SS, et al. Defining the optimal selenium dose for prostate cancer risk reduction: insights from the U-shaped relationship between selenium status, DNA damage, and apoptosis. Dose Response. 2009;8(3):285-300. (PubMed)
166. Ravn-Haren G, Krath BN, Overvad K, et al. Effect of long-term selenium yeast intervention on activity and gene expression of antioxidant and xenobiotic metabolising enzymes in healthy elderly volunteers from the Danish Prevention of Cancer by Intervention by Selenium (PRECISE) pilot study. Br J Nutr. 2008;99(6):1190-1198. (PubMed)