If you appreciate this website, please help us maintain and update the Micronutrient Information Center. Make a gift during Oregon State University's annual giving drive.
April 24 is Dam Proud Day, our annual day of giving. Support the Linus Pauling Institute with a donation on this day and help us unlock over $60,000 of challenge gifts!
Learn more on our Dam Proud Day website
Contents

Vitamin K functions as a cofactor for the enzyme, γ-glutamylcarboxylase, which catalyzes the carboxylation of the amino acid glutamic acid (Glu) to γ-carboxyglutamic acid (Gla). Vitamin K-dependent γ-carboxylation that occurs only on specific glutamic acid residues in identified vitamin K-dependent proteins (VKDP) is critical for their ability to bind calcium (5).
Although vitamin K is a fat-soluble vitamin, the body stores very small amounts that are rapidly depleted without regular dietary intake. Perhaps because of its limited ability to store vitamin K, the body recycles it through a process called the vitamin K-epoxide cycle (Figure 2). The vitamin K cycle allows a small amount of vitamin K to be reused many times for protein carboxylation, thus decreasing the dietary requirement. Briefly, vitamin K hydroquinone (reduced form) is oxidized to vitamin K epoxide (oxidized form). The reaction enables γ-glutamylcarboxylase to carboxylate selective glutamic acid residues on vitamin K-dependent proteins. The recycling of vitamin K epoxide (oxidized form) to hydroquinone (reduced form) is carried out by two reactions that reduce vitamin K epoxide (KO) to vitamin K quinone and then to vitamin K hydroquinone (KH2; Figure 2). Additionally, the enzyme vitamin K-epoxide reductase (VKOR) catalyzes the reduction of KO to vitamin K quinone and may be involved — as well as another yet-to-defined reductase — in the production of KH2 from vitamin K quinone (6, 7). The anticoagulant drug warfarin acts as a vitamin K antagonist by inhibiting VKOR activity, hence preventing vitamin K recycling (see Coagulation).

The ability to bind calcium ions (Ca2+) is required for the activation of the several vitamin K-dependent clotting factors, or proteins, in the coagulation (clotting) cascade. The term ‘coagulation cascade’ refers to a series of events, each dependent on the other, that stops bleeding through clot formation. Vitamin K-dependent γ-carboxylation of specific glutamic acid residues in those proteins makes it possible for them to bind calcium. Factors II (prothrombin), VII, IX, and X make up the core of the coagulation cascade. Protein Z appears to enhance the action of thrombin (the activated form of prothrombin) by promoting its association with phospholipids in cell membranes. Protein C and protein S are anticoagulant proteins that provide control and balance in the coagulation cascade; protein Z also has an anticoagulatory function. Control mechanisms for the coagulation cascade exist since uncontrolled clotting may be as life threatening as uncontrolled bleeding. Vitamin K-dependent coagulation factors are synthesized in the liver. Consequently, severe liver disease results in lower blood levels of vitamin K-dependent clotting factors and an increased risk for uncontrolled bleeding (hemorrhage) (8).
Some people are at increased risk of forming clots, which could block the flow of blood in arteries of the heart, brain, or lungs, resulting in myocardial infarction (heart attack), stroke, or pulmonary embolism, respectively. Abnormal clotting is not related to excessive vitamin K intake, and there is no known toxicity associated with vitamin K1 or vitamin K2 (see Toxicity). Some oral anticoagulants, such as warfarin (Jantoven, formerly known as Coumadin), inhibit coagulation by antagonizing the action of vitamin K. Warfarin prevents the recycling of vitamin K by blocking VKOR activity, thus creating a functional vitamin K deficiency (Figure 2). Inadequate γ-carboxylation of vitamin K-dependent coagulation proteins interferes with the coagulation cascade, which inhibits blood clot formation. Large quantities of dietary or supplemental vitamin K can overcome the anticoagulant effect of vitamin K antagonists; thus, patients taking these drugs are cautioned against consuming very large or highly variable quantities of vitamin K (see Drug interactions). Experts now advise a reasonably constant dietary intake of vitamin K that meets current dietary recommendations (90-120 μg/day) for patients taking vitamin K antagonists like warfarin (9, 10).
Vitamin K-dependent γ-carboxylation is essential to several bone-related proteins, including osteocalcin, anticoagulation factor protein S, matrix γ-carboxylated glutamate (Gla) protein (MGP), Gla-rich protein (GRP), and periostin (originally called osteoblast-specific factor-2). Osteocalcin (also known as bone Gla protein) is synthesized by osteoblasts (bone-forming cells); the synthesis of osteocalcin is regulated by the active form of vitamin D, 1,25-dihydroxyvitamin D (calcitriol). The calcium-binding capacity of osteocalcin requires vitamin K-dependent γ-carboxylation of three glutamic acid residues. Although its function in bone mineralization is not fully understood, osteocalcin is required for the growth and maturation of calcium hydroxyapatite crystals (see Osteoporosis) (11).
Protein S appears to play a role in the breakdown of bone mediated by osteoclasts. Individuals with inherited protein S deficiency suffer complications related to increased blood clotting, as well as osteonecrosis (12, 13). Protein S can bind and activate receptors of the TAM family that are involved in phagocytosis. Mutations in TAM receptors can result in visual impairment, defective spermatogenesis, autoimmune disorders, and platelet disorders (14).
MGP has been found in cartilage, bone, and soft tissue, including blood vessel walls, where it is synthesized and secreted by vascular smooth muscle cells. MGP is involved in the inhibition of calcification at various sites, including cartilage, vessel wall, skin elastic fibers, and the trabecular meshwork in the eye (see Vascular calcification) (15, 16). Moreover, several VKDPs, including MGP, have been associated with calcification sites in arteries, skin, kidneys, and eyes in certain inherited conditions, such as pseudoxanthoma elasticum and beta-thalassemia (17, 18).
The vitamin K-dependent proteins, GRP and periostin, are also synthesized in bone tissue, but their roles in bone metabolism are still unclear (19, 20). Expressed in normal human skin and vascular tissues, GRP has been colocalized with abnormal mineral deposits in the extracellular matrix in calcified arteries and calcified skin lesions (21). Expressed in most connective tissues, including skin and bone, periostin was initially associated with cell adhesion and migration. This VKDP also appears to promote angiogenesis (formation of new blood vessels) during cardiac valve degeneration and tumor growth (22, 23).
Current research suggests that reduced γ-glutamylcarboxylase activity and/or lower vitamin K bioavailability may impair the activity of VKDPs and contribute to bone mineralization defects and abnormal soft tissue calcification (see Disease Prevention) (24).
Growth arrest-specific gene 6 protein (Gas6) is a vitamin K-dependent protein that was identified in 1993. It has been found throughout the nervous system, as well in the heart, lungs, stomach, kidneys, and cartilage. Identified as a ligand of the TAM family of transmembrane tyrosine kinase receptors, Gas6 appears to be a cellular growth regulation factor with cell-signaling activities. Gas6 has been involved in diverse cellular functions, including phagocytosis, cell adhesion, cell proliferation, and protection against apoptosis (5). It may also play important roles in the developing and aging nervous system (reviewed in 25). Further, Gas6 appears to regulate platelet signaling and vascular hemostasis (26). Expressed in most tissues and involved in many cellular functions, Gas6 has also been linked to several pathological conditions, including clot formation (thrombogenesis), atherosclerosis, chronic inflammation, and cancer growth (27-29).
Overt vitamin K deficiency results in impaired blood clotting, usually demonstrated by laboratory tests that measure clotting time. Symptoms include easy bruising and bleeding that may be manifested as nosebleeds, bleeding gums, blood in the urine, blood in the stool, tarry black stools, or extremely heavy menstrual bleeding. In infants, vitamin K deficiency may result in life-threatening bleeding within the skull (intracranial hemorrhage) (8).
Multiple biomarkers of vitamin K status exist (reviewed in 30), but only impaired blood coagulation is used as a measure of clinical vitamin K deficiency.
Vitamin K deficiency is uncommon in healthy adults for a number of reasons: (1) vitamin K is widespread in foods (see Food sources); (2) the vitamin K cycle conserves vitamin K (see Vitamin K oxidation-reduction cycle); and (3) bacteria that normally inhabit the large intestine synthesize menaquinones (vitamin K2), although it is unclear whether significant amounts are absorbed and utilized (see Food sources). Adults at risk for vitamin K deficiency include those taking vitamin K antagonists and individuals with significant liver damage or disease (8). Additionally, individuals with fat malabsorption disorders, including inflammatory bowel disease and cystic fibrosis, may be at increased risk of vitamin K deficiency (31-34).
Newborn babies who are exclusively breast-fed are at increased risk for vitamin K deficiency because human milk is relatively low in vitamin K compared to formula. Newborn infants, in general, have low vitamin K status for the following reasons: (1) vitamin K transport across the placental barrier is limited; (2) liver storage of vitamin K is very low; (3) the vitamin K cycle may not be fully functional in newborns, especially premature infants; (4) the vitamin K content of breast milk is low, and immature gut flora (5, 35). Infants whose mothers are on anticonvulsant medication to prevent seizures are also at risk for vitamin K deficiency. Vitamin K deficiency in newborns may result in a bleeding disorder called vitamin K deficiency bleeding (VKDB) of the newborn (reviewed in 36). Because VKDB is life threatening and easily prevented, the American Academy of Pediatrics and a number of similar international organizations recommend that an intramuscular dose of phylloquinone (vitamin K1) be administered to all newborns shortly after birth (reviewed in 37).
Vitamin K and childhood leukemia: In the early 1990s, two retrospective studies were published suggesting a possible association between phylloquinone injections in newborns and the development of childhood leukemia and other forms of childhood cancer. However, two large retrospective studies in the US and Sweden, which reviewed the medical records of 54,000 and 1.3 million children, respectively, found no evidence of a relationship between childhood cancers and phylloquinone injections at birth (38, 39). Moreover, a pooled analysis of six case-control studies, including 2,431 children diagnosed with childhood cancer and 6,338 cancer-free children, found no evidence that phylloquinone injections for newborns increased the risk of childhood leukemia (40). In a policy statement, the American Academy of Pediatrics recommended that routine vitamin K prophylaxis for newborns be continued because VKDB is life threatening and the risks of cancer are unproven and unlikely (41). In the last decade, physicians have reported a rise in late-onset cases of VKDB due to an increasing trend of parental omission or refusal of newborn vitamin K prophylaxis (42, 43).
Lower doses of vitamin K1 for premature infants: The results of two studies of vitamin K levels in premature infants suggest that the standard initial dose of phylloquinone (vitamin K1) for full-term infants (1.0 mg) may be too high for premature infants (44, 45). These findings have led some experts to suggest the use of an initial phylloquinone dose of 0.3 mg/kg for infants with birth weights of less than 1,000 g (2 lbs, 3 oz), and an initial phylloquinone dose of 0.5 mg would probably prevent hemorrhagic disease in newborns (44). Yet, additional studies are needed to determine the best vitamin K prophylaxis in premature infants (46).
In 2001, the US Food and Nutrition Board (FNB) of the Institute of Medicine (now the National Academy of Medicine) established the adequate intake (AI) level for vitamin K based on phylloquinone intake levels in healthy individuals (Table 1). The AI for infants was based on estimated intake of vitamin K from breast milk (47). The FNB did not consider menaquinone intakes; data on menaquinone consumption are limited (48).
The discovery of vitamin K-dependent proteins in bone has led to research on the role of vitamin K in maintaining bone health.
Vitamin K1: Observational studies have found a relationship between phylloquinone (vitamin K1) and age-related bone loss (osteoporosis). The Nurses’ Health Study followed more than 72,000 women for 10 years. In an analysis of this cohort, women whose phylloquinone intakes were lower than 109 micrograms/day (μg/day) had a 30% higher risk for hip fracture compared to women with intakes equal to or above 109 μg/day (49). Another prospective study in over 800 elderly men and women, followed in the Framingham Heart Study for seven years, found that participants with dietary vitamin K intakes in the highest quartile (median, 254 μg/day) had a 65% lower risk of hip fracture than those with intakes in the lowest quartile (median, 56 μg/day) (50). Osteoporotic fractures are often linked to a reduction in bone mineralization. Yet, the investigators found no association between dietary phylloquinone intake and bone mineral density (BMD) in the Framingham subjects (50). While other studies failed to observe associations between dietary phylloquinone intake and measures of bone strength, BMD, or fracture incidence (51, 52), the cross-sectional study of a cohort of 3,199 middle-aged women found that subjects in the highest quartile of dietary phylloquinone intake had significantly greater hip and lumbar spine BMD than those in the lowest quartile (162 μg/day vs. 59 μg/day) (53). Moreover, cross-sectional and case-control studies have reported associations between higher phylloquinone intakes and lower incidence of hip fracture (54, 55).
However, because green leafy vegetables are the primary dietary source of phylloquinone and because they are usually part of a balanced diet, high phylloquinone consumption may simply be an indicator of healthy eating habits, which may — rather than phylloquinone itself — account for all or part of the association reported in observational studies (56).
The few studies that measured plasma phylloquinone generally found that higher circulating levels were associated with lower fracture risk (15, 57, 58). For example, the incidence of vertebral fractures was inversely correlated with lumbar BMD and plasma phylloquinone in a four-year prospective study that included 379 Japanese women ages 30-88 years (57). Yet, observational studies are not designed to making causal inferences, and only randomized controlled trials can evaluate whether phylloquinone may have beneficial effects on bone health (see Vitamin K supplementation studies and osteoporosis).
Vitamin K2: There are few studies on associations between menaquinones (vitamin K2) and bone health, perhaps because of the limited number of dietary sources of menaquinone-4 (MK-4), the main form of vitamin K2 present in Western diets. The Japanese food natto, made of cooked soybeans fermented by Bacillus subtilis natto, is rich in MK-7. In a prospective cohort study that followed 944 Japanese women (ages 20-79 years), total hip BMD at baseline was positively associated with natto intake in postmenopausal women (59). During the three-year follow-up period, the rate of BMD loss at the femoral neck was significantly lower in women consuming natto (>200 μg/day of MK-7) compared to non-consumers. No association was found between natto intake and BMD in premenopausal women (59).
Total hip and femoral neck BMD was also reportedly higher in nearly 2,000 Japanese men aged 65 years and older who regularly consumed at least of one pack per day of natto (≥350 μg/day of MK-7) compared to those consuming less than one pack per week (<50 μg/day of MK-7) (60). Yet, increasing natto consumption also maximizes the intake of other dietary compounds (e.g., soy isoflavones) that have potential benefits for skeletal health; thus, there is need to find reliable measures of vitamin K status. To date, observational studies have failed to unequivocally support an association between circulating menaquinone (MK-7 and MK-4) levels and fracture risk (15, 61).
A meta-analysis that pooled the results of four prospective cohort studies and one nested case-control study found higher total dietary vitamin K intakes to be associated with a lower risk of total fracture (RR, 0.78; 95% CI: 0.56, 0.99) (62). In a recent Japanese cohort study not included in this meta-analysis (i.e., the Murakami Cohort Study), a protective association of dietary vitamin K intake was seen for vertebral fractures (p=0.005); vitamin K intake was not associated with total fracture in women or with either vertebral or total fracture in men (63).
Biomarker of vitamin K status and bone health
Total circulating levels of the bone protein, osteocalcin (OC), have been shown to be sensitive markers of bone formation. Several hormones and growth factors, including vitamin D but not vitamin K, regulate osteocalcin synthesis by osteoblasts. However, vitamin K is an essential cofactor for the γ-carboxylation of three glutamic acid residues in osteocalcin. Undercarboxylation of osteocalcin in human bone and serum has been linked to poor vitamin K status. The degree of osteocalcin γ-carboxylation is responsive to vitamin K nutritional interventions, and thus is used as a relative indicator of vitamin K status (11).
Circulating levels of uncarboxylated osteocalcin (ucOC) were found to be higher in postmenopausal women than premenopausal women and markedly higher in women over the age of 70. Also, high ratios of ucOC to total OC (ucOC/OC) appear to be predictive of hip fracture risk in elderly women (64, 65). Although vitamin K deficiency would seem the most likely cause of elevated blood ucOC/OC ratio, some investigators have documented an inverse relationship between biochemical measures of vitamin D nutritional status and ucOC levels, as well as a significant lowering of ucOC/OC ratio by vitamin D supplementation (66). It has been suggested that increased circulating ucOC/OC ratio could reflect a poor overall nutritional status that would include vitamin D inadequacy, which would explain the above-mentioned observations (67). However, in several randomized, placebo-controlled intervention studies conducted in young girls (67, 68) and postmenopausal women (69), vitamin D supplementation failed to decrease ucOC/OC ratios or show any additive effect on ucOC/OC lowering by supplemental vitamin K.
Vitamin K1 supplementation: The review of five randomized clinical trials that assessed the effect of phylloquinone (vitamin K1) supplementation on hip BMD using doses ranging from 200 μg/day to 5,000 μg/day for durations of 12 to 36 months found little promising benefit for bone health (15). Although supplementation with phylloquinone decreased ucOC levels in all five studies, only one study reported an effect of supplemental phylloquinone on BMD (70). In this study, 150 postmenopausal women were randomized to receive a placebo, minerals (500 mg/day of calcium, 150 mg/day of magnesium, and 10 mg/day of zinc) plus vitamin D (320 IU/day), or minerals, vitamin D, and phylloquinone (1,000 μg/day). The rate of BMD loss at the femoral neck, but not at the lumbar spine, was significantly lower in subjects with supplemental phylloquinone compared to the other two groups. Thus, evidence of a putative benefit of phylloquinone on bone health in older adults is considered weak. None of the studies were designed to assess the effect of phylloquinone on osteoporotic-related fractures. Further investigation may seek to evaluate whether phylloquinone supplementation could improve skeletal health in subjects at high-risk for vitamin K inadequacy (e.g., individuals with malabsorption syndromes or cystic fibrosis).
Vitamin K2 supplementation: Pharmacological doses of menaquinone-4 (MK-4; brand name, menatetrenone) are currently used in Japan in the treatment of osteoporosis (71). Accordingly, most intervention trials investigating the effect of high-dose MK-4 on bone loss have been conducted in Japanese postmenopausal women. In a three-year placebo-controlled trial among postmenopausal women with osteopenia, adding a MK-7 supplement (375 μg/day) to combined calcium-vitamin D supplementation did not affect BMD or other bone health parameters despite reductions in serum ucOC (72). At present, the potential role for supplemental menaquinones on bone health still needs to be established in large, randomized, and well-controlled trials.
A 2019 systematic review and meta-analysis of clinical trials in postmenopausal women or women with osteoporosis found vitamin K supplementation — of any form — lowered risk of clinical fracture compared to controls (OR, 0.72; 95% CI, 0.55-0.95; 9 trials varying from 6 months to 2 years: 3 using phylloquinone, 5 using MK-4, and 1 using MK-7) (73). However, no benefit of vitamin K supplementation was seen for vertebral fractures (7 trials) or BMD (22 trials). The authors of this meta-analysis noted that the high heterogeneity of the included trials (i.e., form and dose of vitamin K, use of other supplements and drugs by participants, and treatment length) makes it difficult to inform clinical recommendations (73).
Certain oral anticoagulants, such as warfarin, are known to be antagonists of vitamin K (see Coagulation). Few studies have examined chronic use of warfarin and risk of fracture in older women. One study reported no association between long-term warfarin treatment and fracture risk (74), while another one found a significantly higher risk of rib and vertebral fractures in warfarin users compared to nonusers (75). Additionally, a study in elderly patients with atrial fibrillation reported that long-term warfarin treatment was associated with a significantly higher risk of osteoporotic fracture in men but not in women (76). A meta-analysis of the results of 11 published studies found that oral anticoagulation therapy was associated with a very modest reduction in BMD at the wrist and no change in BMD at the hip or spine (77). The development of new anticoagulants that do not block vitamin K recycling may offer a safer alternative to the use of vitamin K antagonists (78).
Observational studies examining vitamin K intake in relation to cardiovascular-related mortality have found conflicting results (48). An inverse relationship between vitamin K intake and mortality was reported in a US national survey (NHANES III) of 3,401 participants (79). Adequate vs. inadequate vitamin K intakes (based on sex-specific AI: 90 μg/day for women and 120 μg/day for men) were associated with a 22% lower risk of cardiovascular disease (CVD)-related mortality and a 15% lower risk of all-cause mortality. The report also indicated that, while over two-thirds of individuals with chronic kidney disease had vitamin K intakes below the AI, the risk of CVD mortality was 41% lower in those with adequate compared to suboptimal intakes (79). However, higher vitamin K intakes were not associated with lower CVD mortality in a prospective study that followed 7,216 older adults at risk for developing CVD (80). This study associated higher intakes of phylloquinone, but not of menaquinones, with lower risk of all-cause mortality. More recently, in a prospective study that followed a cohort of 33,289 Dutch men and women for an average of 16.8 years (the EPIC-Netherlands), neither phylloquinone intake nor menaquinone intake at baseline was associated with mortality from cardiovascular disease, coronary heart disease, stroke, or all causes (81). Yet, a prospective study of 56,048 men and women participating in the Danish Diet, Cancer, and Health cohort reported higher phylloquinone intakes to be associated with lower risks of both cardiovascular disease-related mortality and all-cause mortality (82). Moreover, a few studies have examined the association of fasting circulating phylloquinone with cardiovascular-related or all-cause mortality. In a recent meta-analysis of individual participant data from three prospective cohort studies (the Multi-Ethnic Study of Atherosclerosis; the Health, Aging, and Body Composition Study, and the Framingham Offspring Study), a blood phylloquinone concentration less than or equal to 0.5 nmol/L (n=1,081) was associated with a 19% higher risk of all-cause mortality compared to a blood concentration >1.0 nmol/L (n=1,698; 83). This analysis did not find circulating phylloquinone to be linked with incident cardiovascular disease (83).
Observational studies offer limited support of an inverse relationship between phylloquinone intake and risk of incident cardiovascular disease, despite high intakes being sometimes regarded as a marker of healthy dietary habits associated with low cardiovascular risk (reviewed in 84). A prospective cohort study of 16,057 Dutch women (ages 49-70 years), followed for a mean period of 8.1 years, found a 9% reduction in risk for coronary heart disease (CHD) per each incremental 10 μg/day increase in menaquinone intake (85). In another earlier Dutch study that examined 4,807 healthy men and women 55 years and older, participants in the highest tertile of menaquinone intake (>32.7 μg/day) had a 41% lower risk of incident CHD and a 26% lower risk of all-cause mortality than those in the lowest tertile (<21.6 μg/day) (86). In addition, menaquinone intake was found to be inversely associated with aortic calcification, a major risk factor for CVD (86). A smaller prospective study among 2,987 Norwegian adults, followed for an average of 10.8 years, reported higher dietary intakes of vitamin K2, but not of vitamin K1, were associated with a lower risk of CHD (87). While these data are interesting, it is important to note that vitamin K2 may solely be a marker of a heart-healthy diet and might not itself be cardioprotective. Most recently, the prospective Danish Diet, Cancer, and Health Study followed 53,372 older adults for an average of 21 years, finding both phylloquinone and menaquinone intake at baseline to be inversely associated with hospitalizations due to atherosclerotic cardiovascular events, including ischemic heart disease, ischemic stroke, and peripheral arterial disease (88).
Large-scale supplementation trials are needed to determine whether vitamin K1 or vitamin K2 reduces the risk of CHD or cardiovascular-related events like myocardial infarction or stroke.
One of the hallmarks of cardiovascular disease is the presence of atherosclerotic plaques in arterial walls. Plaque rupture that causes blood clot formation (thrombogenesis) is the usual cause of a myocardial infarction (heart attack) or stroke. While calcification of the plaques occurs as the atherosclerosis progresses, it is unclear whether calcification increases plaque instability and could predict risk of rupture and thrombogenesis (89). However, calcification may be predictive of future cardiovascular events, especially in those with chronic kidney disease (90). A meta-analysis of 30 prospective cohort studies, including a total of 218,080 participants, found that the presence of vascular calcification was associated with an overall three- to four-fold increased risk of cardiovascular events and mortality (91). An early population-based study of postmenopausal women (ages, 60-79 years) observed that the younger women (60-69 years) with aortic calcifications had lower vitamin K intakes than those without aortic calcifications, but this was not true for older women (70-79 years) (92). A prospective cohort study in 807 men and women, 39-45 years of age, did not find any correlation between dietary phylloquinone intake and coronary artery calcification, as measured non-invasively by computed tomography (93). Additionally, neither phylloquinone nor menaquinone intake was associated with calcification of breast arteries in a cross-sectional study of 1,689 women ages 49-70 years (94). However, in another cross-sectional study, the upper vs. lowest quartile of menaquinone (MK-4 to MK-10) intake (median intakes, 48.5 μg/day vs. 18 μg/day) was found to be associated with a 20% reduced prevalence of coronary artery calcification in 564 postmenopausal women (95).
Research has uncovered possible mechanisms by which vitamin K may inhibit mineralization (calcification) of vessels while promoting bone mineralization. The potential mechanisms, although not yet fully understood, implicate vitamin K-dependent proteins, including matrix Gla protein (MGP), Gla-rich protein (GRP), and Gas6 (96-98). Secreted by various cell types, such as vascular smooth muscle cells (VSMCs) in arterial vessel walls, MGP appears to be important for the prevention of calcification of soft tissues, including cartilage, vasculature, skin, and trabecular meshwork cells in the eye (15, 99). In MGP knockout mice, conversion of VSMCs into bone-like cells and extensive vessel calcification results in large vessel rupture and premature death. In humans, defective MGP gene has been linked to Keutel syndrome, a rare inherited condition characterized in particular by abnormal cartilage calcification and pulmonary artery stenosis (narrowing). Calcification prevention by MGP involved several mechanisms, including the binding to calcium crystals and the inhibition of proteins (bone morphogenic proteins; BMPs) known to promote ectopic bone formation (reviewed in 100).
Calcium-binding activity of MGP is regulated by two types of modifications (known as post-translational modifications since they take place after protein synthesis): the vitamin K-dependent carboxylation of up to five Glu residues and the phosphorylation of serine residues. A variation in the sequence (polymorphism) of the gene for MGP leading to a threonine-to-alanine transition in one of the five Gla domains of the protein may possibly prevent carboxylation and elicit a change in MGP ability to bind calcium. This polymorphism, known as MGPThr83Ala, has been associated with the progression of coronary artery calcification over a mean period of 10.6 years in a community-based prospective study that followed 605 middle-aged men and women (101). This association was only observed among participants without detectable calcification at baseline and not in those who had baseline calcification (101). Interestingly, MGPThr83Ala was also associated with higher risk of myocardial infarction and femoral artery calcification in carriers of the genotype (102).
Additionally, a small study initially found that, while undercarboxylated MGP (ucMGP) was absent from the innermost lining of the carotid arteries in healthy subjects, the majority of MGP in the carotid arterial lining of patients with atherosclerosis was undercarboxylated (103). In another study that examined the association between circulating MGP and incident cardiovascular events in 577 older men and women followed for a mean period of 5.6 years, the risk of cardiovascular disease (i.e., coronary artery disease, peripheral arterial disease, and cerebrovascular disease) was two- to three-fold greater in subjects in the highest vs. lowest tertile of plasma dephosphorylated and undercarboxylated MGP (dp-ucMGP) (104). The results of another prospective study suggested that circulating dp-ucMGP may be predictive of mortality risk in subjects with overt vascular disease (105). Indeed, the risk of cardiovascular-related and all-cause mortality was found to be nearly doubled in subjects with coronary artery disease or stroke in the highest vs. lowest quartile of dp-ucMGP concentrations (105).
Because suboptimal vitamin K nutritional status may limit carboxylation and result in biologically inactive ucMGP, it has been speculated that vitamin K supplementation may protect against vascular calcification. A three-year, double-blind, controlled trial investigated the potential effect of vitamin K on the progression of coronary calcification in 401 older, community-dwelling adults (ages, 60-80 years) free of cardiovascular disease at baseline (106). The participants were randomized to receive a daily multivitamin plus calcium and vitamin D with or without 500 mg of phylloquinone. Using measurements of coronary artery calcification at baseline and follow up, it was found that phylloquinone supplementation was able to limit the progression of vascular calcification and reduce plasma dp-ucMGP compared to control (106, 107). Although circulating dp-ucMGP was correlated to various markers of vitamin K status, no association with measures of coronary artery calcification were found (107). Smaller trials in those at high risk for coronary artery disease (108) or with existing aortic valve calcification (109), type 2 diabetes mellitus (110), or kidney disease (111, 112) have reported no benefit of vitamin K2 supplementation on progression of vascular calcification. A meta-analysis of controlled clinical trials found that vitamin K supplementation reduced dp-ucMGP (7 trials), reduced ucOC (4 trials), and decreased progression of vascular calcification (3 trials) (113). Yet, the authors emphasize that any conclusions from their pooled analysis are limited by the high heterogeneity of the trials with respect to the form and dose of vitamin K utilized, as well as the assays that assessed vascular calcification (113).
Thus, further investigations are necessary to examine the role of other vitamin K-dependent proteins (e.g., GRP, periostin, Gas6) in human atherosclerotic plaque calcification and to evaluate the effect of supplemental vitamin K on the progression of vascular calcification and risk of cardiovascular disease.
Several cross-sectional studies have reported increased vascular calcium scores (a means to quantify vascular calcification) in chronic users of vitamin K antagonists compared to nonusers (reviewed in 114). Warfarin therapy has also been associated with higher circulating concentrations of dp-ucMGP in a prospective study that examined vascular calcification in subjects with cardiovascular disease (105). Newly developed direct inhibitors of coagulation factors that do not interfere with VKDP activity may be more suitable than vitamin K antagonists, especially with regards to vascular calcification (114).
Osteoarthritis, a degenerative joint condition that affects more than 32 million US adults (115), is characterized by the breakdown of articular cartilage (i.e., cartilage within the joint). Because several vitamin K-dependent proteins are present in cartilage and in bone (116), vitamin K inadequacy may have a role in the development of osteoarthritis. A few observational studies have investigated a possible link between vitamin K intake or status and osteoarthritis. A cross-sectional study among 719 Japanese older adults found dietary intake of vitamin K to be inversely associated with knee osteoarthritis (117). In the Framingham Offspring Study (n=672; mean age, 66 years), higher plasma concentrations of phylloquinone were associated with a lower risk of hand, but not knee, osteoarthritis (118). A longitudinal study of 1,180 US adults (mean age, 62 years) found that low plasma concentrations of phylloquinone (≤0.5 nM) at baseline — indicative of a subclinical vitamin K deficiency — were associated with a 56% increase in risk of knee osteoarthritis after 30 months compared to those with higher plasma concentrations (119). In a more recent longitudinal study among 523 older US adults participating in the Health, Aging, and Body Composition Study, those with extremely low plasma concentrations of phylloquinone (<0.2 nM) at baseline had increased progression of knee osteoarthritis over three years, assessed by MRI of articular cartilage and the meniscus; those with higher plasma concentrations experienced no significant progression of knee osteoarthritis (120). Moreover, recent studies have associated use of the vitamin K antagonist drugs with higher risks of osteoarthritis (121) and joint replacement (121) of the knee and hip compared to nonusers.
While these observational data are interesting, randomized controlled trials are needed to determine whether vitamin K supplementation in those with low vitamin K status might help prevent or treat osteoarthritis. In an ancillary study of a double-blind, controlled trial examining the effects of vitamin K supplementation on bone loss and vascular calcification in older adults, no effects of phylloquinone supplementation (500 μg/day) were found on incidence of hand osteoarthritis after three years (122). Study participants were not screened for vitamin K status, and in a subgroup analysis, those with serum phylloquinone ≤1 nM at baseline that reached >1 nM at year 3 had less joint deterioration. These data infer that only those individuals with low vitamin K status benefit from vitamin K supplementation. Unfortunately, no measures were available for knee osteoarthritis, and it is well established that hand and knee osteoarthritis represent different phenotypes. Additional clinical trials that specifically examine the effect of vitamin K supplementation on osteoarthritis development are needed, especially in those with inadequate vitamin K status.
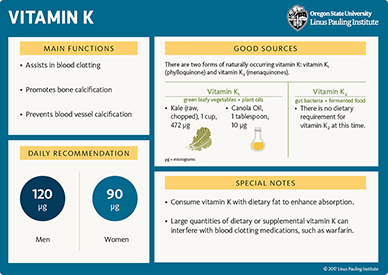
A US national survey, NHANES 2011-2012, found that average dietary intakes of vitamin K (all forms) vary greatly among individuals, with values ranging from 80 to 195 μg/day of phylloquinone for men and 78 to 223 μg/day of phylloquinone for women (123). Mean intakes for both men and women were 117 μg/day of phylloquinone; however, 57% of men and 37% of women did not meet the Adequate Intake (AI) level (123).
Phylloquinone (vitamin K1) is the major dietary form of vitamin K in most diets. Green leafy vegetables and some plant oils (soybean, canola, olive, and cottonseed) are major contributors of dietary vitamin K. Mixed dishes have also been found to significantly contribute to vitamin K intake in the US (123). However, phylloquinone bioavailability from green vegetables is lower than from oil or supplements. Also, the phylloquinone content of green vegetables depends on their content in chlorophyll (green pigment), so that outer leaves have more phylloquinone than inner leaves. The efficiency of phylloquinone intestinal absorption varies among plant sources and is increased with the addition of a fat source to a meal. Finally, the hydrogenation of vegetable oils may decrease the absorption and biological effect of dietary phylloquinone (reviewed in 2, 9). If you wish to check foods for their nutrient content, including phylloquinone, search USDA’s FoodData Central. A number of phylloquinone-rich foods are listed in Table 2, with their content in phylloquinone expressed in micrograms (μg).
Menaquinones (vitamin K2) are primarily of microbial origins and thus commonly found in fermented foods, such as cheese, curds, and natto (fermented soybeans) (124, 125). MK-4 is the only menaquinone that is not produced by bacteria. MK-4 is formed from menadione (a synthetic form of vitamin K) found in animal feeds or is converted in a tissue-specific way from multiple dietary forms of vitamin K, including phylloquinone and various menaquinones (4, 126). Menaquinone-4 is found in dairy products, including milk, and in some meats (125). Longer chain menaquinones are found in limited fermented food products. The Japanese fermented soybean-based, natto, is rich in MK-7 (998 μg/100 g) and also contains MK-8 (84 μg/100 g). Some cheeses contain MK-8 and MK-9 (2, 125). Additionally, animal livers are a source of long-chain menaquinones (MK-7 to MK-13) (9).
Food composition databases, including USDA’s FoodData Central, have limited data on menaquinone content in foods. Thus, menaquinone contribution to total vitamin K intakes is difficult to estimate and likely to vary between populations with different food consumption practices (2, 125). Bacteria that normally colonize the large intestine (colon) can synthesize menaquinones. It was initially thought that up to 50% of the human vitamin K requirement might be met by bacterial synthesis. However, all forms of vitamin K are absorbed in the small intestine via a mechanism requiring bile salts, while most of the menaquinone production takes place in the colon where bile salts are lacking. Current research suggests that the contribution of bacterial synthesis to vitamin K status is much less than previously thought, although the exact contribution remains unclear (16, 127).
In the US, both phylloquinone and menaquinones are available without a prescription in multivitamin, single-nutrient, or multiple-nutrient supplements in varying doses; vitamin K content of multivitamins typically range from 20 to 120 μg per tablet (128). The US Food and Drug Administration (FDA) has not authorized any health claims for any forms of vitamin K.
Although allergic reactions are possible, there is no known toxicity associated with high doses (dietary or supplemental) of the phylloquinone (vitamin K1) or menaquinone (vitamin K2) forms of vitamin K (47). The same is not true for synthetic menadione (vitamin K3) and its derivatives. Menadione can interfere with the function of glutathione, one of the body's natural antioxidants, resulting in oxidative damage to cell membranes. Menadione given by injection has induced liver toxicity, jaundice, and hemolytic anemia (due to the rupture of red blood cells) in infants; therefore, menadione is no longer used for treatment of vitamin K deficiency (5). No tolerable upper intake level (UL) has been established for vitamin K (47).
Large doses of vitamin A and vitamin E have been found to antagonize vitamin K (8). Excess vitamin A appears to interfere with vitamin K absorption, whereas vitamin E may inhibit vitamin K-dependent carboxylase activity and interfere with the coagulation cascade (129). One study in adults with normal coagulation status found that supplementation with 1,000 IU/day of vitamin E for 12 weeks decreased γ-carboxylation of prothrombin, a vitamin K-dependent protein (130). Individuals taking anticoagulatory drugs like warfarin and those who are vitamin K deficient should not take vitamin E supplements without close medical supervision because of the increased risk of hemorrhage (excessive bleeding) (131).
The anticoagulant effect of vitamin K antagonists (e.g., warfarin) may be compromised by very high dietary or supplemental vitamin K intake. Moreover, daily phylloquinone supplements of up to 100 μg are considered safe for patients taking warfarin, but therapeutic anticoagulant stability may be undermined by daily doses of MK-7 as low as 10 to 20 μg (132). It is generally recommended that individuals using warfarin try to consume the AI for vitamin K (90-120 μg/day) and avoid large fluctuations in vitamin K intake that might interfere with the adjustment of their anticoagulant dose (9, 10, 133). The prescription of anti-vitamin K anticoagulants, anticonvulsants (e.g., phenytoin), and anti-tuberculosis drugs (e.g., rifampin and isoniazid) to pregnant or breast-feeding women may place the newborn at increased risk of vitamin K deficiency (134).
The drug amiodarone, used in the management of certain cardiac arrhythmias (irregular heartbeat), including atrial fibrillation, can enhance the anticoagulant effect of warfarin and thus increase the risk of hemorrhage (135, 136). Further, the use of cholesterol-lowering medications (like cholestyramine and colestipol), as well as orlistat, mineral oil, and the fat substitute, olestra, may interfere with fat absorption and affect the absorption of fat-soluble vitamins, including vitamin K (137).
It is not clear whether the AI for vitamin K is enough to optimize the γ-carboxylation of vitamin K-dependent proteins in bone (see Osteoporosis). To consume the amount of vitamin K associated with a decreased risk of hip fracture in the Framingham Heart Study (about 250 μg/day) (50), an individual would need to eat a little more than ½ cup of chopped broccoli or a large salad of mixed greens every day. Though the dietary intake of vitamin K required for optimal function of all vitamin K-dependent proteins is not yet known, the Linus Pauling Institute recommends taking a multivitamin/mineral supplement and eating at least one cup of dark-green leafy vegetables daily. Replacing dietary saturated fats like butter and cheese with monounsaturated fats found in olive oil and canola oil will increase dietary vitamin K intake and may decrease the risk of cardiovascular disease.
Because older adults are at increased risk of osteoporosis and hip fracture, the above recommendation for a multivitamin/mineral supplement and at least one cup of dark green leafy vegetables daily is especially relevant.
Originally written in 2000 by:
Jane Higdon, Ph.D.
Linus Pauling Institute
Oregon State University
Updated in May 2004 by:
Jane Higdon, Ph.D.
Linus Pauling Institute
Oregon State University
Updated in May 2008 by:
Victoria J. Drake, Ph.D.
Linus Pauling Institute
Oregon State University
Updated in July 2014 by:
Barbara Delage, Ph.D.
Linus Pauling Institute
Oregon State University
Updated in May 2022 by:
Victoria J. Drake, Ph.D.
Linus Pauling Institute
Oregon State University
Reviewed in July 2022 by:
Sarah L. Booth, Ph.D.
Director, Vitamin K Research Program
Jean Mayer USDA Human Nutrition Research Center on Aging
Tufts University
Copyright 2000-2024 Linus Pauling Institute
1. Brody T. Nutritional Biochemistry. 2nd ed. San Diego: Academic Press; 1999.
2. Booth SL. Vitamin K: food composition and dietary intakes. Food Nutr Res. 2012;56. (PubMed)
3. Kidd PM. Vitamins D and K as pleiotropic nutrients: clinical importance to the skeletal and cardiovascular systems and preliminary evidence for synergy. Altern Med Rev. 2010;15(3):199-222. (PubMed)
4. Nakagawa K, Hirota Y, Sawada N, et al. Identification of UBIAD1 as a novel human menaquinone-4 biosynthetic enzyme. Nature. 2010;468(7320):117-121. (PubMed)
5. Ferland G. Vitamin K. In: ISLI, ed. Present Knowledge in Nutrition. 10th ed: John Wiley & Sons; 2012:230-247.
6. Rishavy MA, Hallgren KW, Wilson LA, Usubalieva A, Runge KW, Berkner KL. The vitamin K oxidoreductase is a multimer that efficiently reduces vitamin K epoxide to hydroquinone to allow vitamin K-dependent protein carboxylation. J Biol Chem. 2013;288(44):31556-31566. (PubMed)
7. Tie JK, Jin DY, Straight DL, Stafford DW. Functional study of the vitamin K cycle in mammalian cells. Blood. 2011;117(10):2967-2974. (PubMed)
8. Olson RE. Vitamin K. In: Shils ME, Olson JA, Shike M, Ross AC, eds. Modern Nutrition in Health and Disease. 9th ed. Baltimore: Williams & Wilkins; 1999:363-380.
9. Holmes MV, Hunt BJ, Shearer MJ. The role of dietary vitamin K in the management of oral vitamin K antagonists. Blood Rev. 2012;26(1):1-14. (PubMed)
10. Violi F, GY L, P P, D P. Interaction between dietary vitamin K intake and anticoagulation by vitamin K antagonists: is it really true?: a systematic review. Medicine (Baltimore). 2016;95(10):e2895. (PubMed)
11. Gundberg CM, Lian JB, Booth SL. Vitamin K-dependent carboxylation of osteocalcin: friend or foe? Adv Nutr. 2012;3(2):149-157. (PubMed)
12. Pierre-Jacques H, Glueck CJ, Mont MA, Hungerford DS. Familial heterozygous protein-S deficiency in a patient who had multifocal osteonecrosis. A case report. J Bone Joint Surg Am. 1997;79(7):1079-1084. (PubMed)
13. Rawat RS, Mehta Y, Arora D, Trehan N. Asymptomatic type B right atrial thrombus in a case with protein S deficiency. Ann Card Anaesth. 2014;17(3):237-239. (PubMed)
14. van der Meer JH, van der Poll T, van 't Veer C. TAM receptors, Gas6, and protein S: roles in inflammation and hemostasis. Blood. 2014;123(16):2460-2469. (PubMed)
15. Booth SL. Roles for vitamin K beyond coagulation. Annu Rev Nutr. 2009;29:89-110. (PubMed)
16. Shea MK, Booth SL. Vitamin K. Adv Nutr. 2022;13(1):350-351. (PubMed)
17. Boraldi F, Annovi G, Guerra D, et al. Fibroblast protein profile analysis highlights the role of oxidative stress and vitamin K recycling in the pathogenesis of pseudoxanthoma elasticum. Proteomics Clin Appl. 2009;3(9):1084-1098. (PubMed)
18. Boraldi F, Garcia-Fernandez M, Paolinelli-Devincenzi C, et al. Ectopic calcification in beta-thalassemia patients is associated with increased oxidative stress and lower MGP carboxylation. Biochim Biophys Acta. 2013;1832(12):2077-2084. (PubMed)
19. Coutu DL, Wu JH, Monette A, Rivard GE, Blostein MD, Galipeau J. Periostin, a member of a novel family of vitamin K-dependent proteins, is expressed by mesenchymal stromal cells. J Biol Chem. 2008;283(26):17991-18001. (PubMed)
20. Viegas CS, Simes DC, Laize V, Williamson MK, Price PA, Cancela ML. Gla-rich protein (GRP), a new vitamin K-dependent protein identified from sturgeon cartilage and highly conserved in vertebrates. J Biol Chem. 2008;283(52):36655-36664. (PubMed)
21. Viegas CS, Cavaco S, Neves PL, et al. Gla-rich protein is a novel vitamin K-dependent protein present in serum that accumulates at sites of pathological calcifications. Am J Pathol. 2009;175(6):2288-2298. (PubMed)
22. Hakuno D, Kimura N, Yoshioka M, et al. Periostin advances atherosclerotic and rheumatic cardiac valve degeneration by inducing angiogenesis and MMP production in humans and rodents. J Clin Invest. 2010;120(7):2292-2306. (PubMed)
23. Kudo Y, Siriwardena BS, Hatano H, Ogawa I, Takata T. Periostin: novel diagnostic and therapeutic target for cancer. Histol Histopathol. 2007;22(10):1167-1174. (PubMed)
24. Vanakker OM, Martin L, Schurgers LJ, et al. Low serum vitamin K in PXE results in defective carboxylation of mineralization inhibitors similar to the GGCX mutations in the PXE-like syndrome. Lab Invest. 2010;90(6):895-905. (PubMed)
25. Ferland G. Vitamin K and the nervous system: an overview of its actions. Adv Nutr. 2012;3(2):204-212. (PubMed)
26. Laurance S, Lemarie CA, Blostein MD. Growth arrest-specific gene 6 (gas6) and vascular hemostasis. Adv Nutr. 2012;3(2):196-203. (PubMed)
27. Robins RS, Lemarie CA, Laurance S, Aghourian MN, Wu J, Blostein MD. Vascular Gas6 contributes to thrombogenesis and promotes tissue factor up-regulation after vessel injury in mice. Blood. 2013;121(4):692-699. (PubMed)
28. Rothlin CV, Leighton JA, Ghosh S. Tyro3, Axl, and Mertk receptor signaling in inflammatory bowel disease and colitis-associated cancer. Inflamm Bowel Dis. 2014;20(8):1472-80. (PubMed)
29. Tjwa M, Moons L, Lutgens E. Pleiotropic role of growth arrest-specific gene 6 in atherosclerosis. Curr Opin Lipidol. 2009;20(5):386-392. (PubMed)
30. Card DJ, Gorska R, Harrington DJ. Laboratory assessment of vitamin K status. J Clin Pathol. 2020;73(2):70-75. (PubMed)
31. Jagannath VA, Fedorowicz Z, Thaker V, Chang AB. Vitamin K supplementation for cystic fibrosis. Cochrane Database Syst Rev. 2013;4:CD008482. (PubMed)
32. Nakajima S, Iijima H, Egawa S, et al. Association of vitamin K deficiency with bone metabolism and clinical disease activity in inflammatory bowel disease. Nutrition. 2011;27(10):1023-1028. (PubMed)
33. Nowak JK, Grzybowska-Chlebowczyk U, Landowski P, et al. Prevalence and correlates of vitamin K deficiency in children with inflammatory bowel disease. Sci Rep. 2014;4:4768. (PubMed)
34. Dong R, Wang N, Yang Y, et al. Review on vitamin K deficiency and its biomarkers: focus on the novel application of PIVKA-II in clinical practice. Clin Lab. 2018;64(4):413-424. (PubMed)
35. Araki S, Shirahata A. Vitamin K deficiency bleeding in infancy. Nutrients. 2020;12(3):780. (PubMed)
36. Shearer MJ. Vitamin K deficiency bleeding (VKDB) in early infancy. Blood Rev. 2009;23(2):49-59. (PubMed)
37. Jullien S. Vitamin K prophylaxis in newborns. BMC Pediatr. 2021;21(Suppl 1):350. (PubMed)
38. Klebanoff MA, Read JS, Mills JL, Shiono PH. The risk of childhood cancer after neonatal exposure to vitamin K. N Engl J Med. 1993;329(13):905-908. (PubMed)
39. Ekelund H, Finnstrom O, Gunnarskog J, Kallen B, Larsson Y. Administration of vitamin K to newborn infants and childhood cancer. BMJ. 1993;307(6896):89-91. (PubMed)
40. Roman E, Fear NT, Ansell P, et al. Vitamin K and childhood cancer: analysis of individual patient data from six case-control studies. Br J Cancer. 2002;86(1):63-69. (PubMed)
41. American Academy of Pediatrics Committee on Fetus and Newborn. Controversies concerning vitamin K and the newborn. Pediatrics. 2003;112(1 Pt 1):191-192. (PubMed)
42. Schulte R, Jordan LC, Morad A, Naftel RP, Wellons JC, 3rd, Sidonio R. Rise in late onset vitamin K deficiency bleeding in young infants because of omission or refusal of prophylaxis at birth. Pediatr Neurol. 2014;50(6):564-568. (PubMed)
43. Majid A, Blackwell M, Broadbent RS, et al. Newborn vitamin K prophylaxis: a historical perspective to understand modern barriers to uptake. Hosp Pediatr. 2019;9(1):55-60. (PubMed)
44. Costakos DT, Greer FR, Love LA, Dahlen LR, Suttie JW. Vitamin K prophylaxis for premature infants: 1 mg versus 0.5 mg. Am J Perinatol. 2003;20(8):485-490. (PubMed)
45. Kumar D, Greer FR, Super DM, Suttie JW, Moore JJ. Vitamin K status of premature infants: implications for current recommendations. Pediatrics. 2001;108(5):1117-1122. (PubMed)
46. Ardell S, Offringa M, Ovelman C, Soll R. Prophylactic vitamin K for the prevention of vitamin K deficiency bleeding in preterm neonates. Cochrane Database Syst Rev. 2018;2:CD008342. (PubMed)
47. Food and Nutrition Board, Institute of Medicine. Vitamin K. Dietary Reference Intakes for Vitamin A, Vitamin K, Arsenic, Boron, Chromium, Copper, Iodine, Iron, Manganese, Molybdenum, Nickel, Silicon, Vanadium, and Zinc. Washington, D.C.: National Academy Press; 2001:162-196. (National Academy Press)
48. Shea MK, Berkner KL, Ferland G, Fu X, Holden RM, Booth SL. Perspective: Evidence before enthusiasm-a critical review of the potential cardiovascular benefits of vitamin K. Adv Nutr. 2021;12(3):632-646. (PubMed)
49. Feskanich D, Weber P, Willett WC, Rockett H, Booth SL, Colditz GA. Vitamin K intake and hip fractures in women: a prospective study. Am J Clin Nutr. 1999;69(1):74-79. (PubMed)
50. Booth SL, Tucker KL, Chen H, et al. Dietary vitamin K intakes are associated with hip fracture but not with bone mineral density in elderly men and women. Am J Clin Nutr. 2000;71(5):1201-1208. (PubMed)
51. Rejnmark L, Vestergaard P, Charles P, et al. No effect of vitamin K1 intake on bone mineral density and fracture risk in perimenopausal women. Osteoporos Int. 2006;17(8):1122-1132. (PubMed)
52. McLean RR, Booth SL, Kiel DP, et al. Association of dietary and biochemical measures of vitamin K with quantitative ultrasound of the heel in men and women. Osteoporos Int. 2006;17(4):600-607. (PubMed)
53. Macdonald HM, McGuigan FE, Lanham-New SA, Fraser WD, Ralston SH, Reid DM. Vitamin K1 intake is associated with higher bone mineral density and reduced bone resorption in early postmenopausal Scottish women: no evidence of gene-nutrient interaction with apolipoprotein E polymorphisms. Am J Clin Nutr. 2008;87(5):1513-1520. (PubMed)
54. Apalset EM, Gjesdal CG, Eide GE, Tell GS. Intake of vitamin K1 and K2 and risk of hip fractures: The Hordaland Health Study. Bone. 2011;49(5):990-995. (PubMed)
55. Torbergsen AC, Watne LO, Wyller TB, et al. Vitamin K1 and 25(OH)D are independently and synergistically associated with a risk for hip fracture in an elderly population: A case control study. Clin Nutr. 2015;34(1):101-106. (PubMed)
56. Booth SL, Mayer J. Warfarin use and fracture risk. Nutr Rev. 2000;58(1):20-22. (PubMed)
57. Tsugawa N, Shiraki M, Suhara Y, et al. Low plasma phylloquinone concentration is associated with high incidence of vertebral fracture in Japanese women. J Bone Miner Metab. 2008;26(1):79-85. (PubMed)
58. Moore AE, Kim E, Dulnoan D, et al. Serum vitamin K1 (phylloquinone) is associated with fracture risk and hip strength in post-menopausal osteoporosis: A cross-sectional study. Bone. 2020;141:115630. (PubMed)
59. Ikeda Y, Iki M, Morita A, et al. Intake of fermented soybeans, natto, is associated with reduced bone loss in postmenopausal women: Japanese Population-Based Osteoporosis (JPOS) Study. J Nutr. 2006;136(5):1323-1328. (PubMed)
60. Fujita Y, Iki M, Tamaki J, et al. Association between vitamin K intake from fermented soybeans, natto, and bone mineral density in elderly Japanese men: the Fujiwara-kyo Osteoporosis Risk in Men (FORMEN) study. Osteoporos Int. 2012;23(2):705-714. (PubMed)
61. Kaneki M, Hodges SJ, Hosoi T, et al. Japanese fermented soybean food as the major determinant of the large geographic difference in circulating levels of vitamin K2: possible implications for hip-fracture risk. Nutrition. 2001;17(4):315-321. (PubMed)
62. Hao G, Zhang B, Gu M, et al. Vitamin K intake and the risk of fractures: A meta-analysis. Medicine (Baltimore). 2017;96(17):e6725. (PubMed)
63. Platonova K, Kitamura K, Watanabe Y, et al. Dietary calcium and vitamin K are associated with osteoporotic fracture risk in middle-aged and elderly Japanese women, but not men: the Murakami Cohort Study. Br J Nutr. 2021;125(3):319-328. (PubMed)
64. Szulc P, Chapuy MC, Meunier PJ, Delmas PD. Serum undercarboxylated osteocalcin is a marker of the risk of hip fracture in elderly women. J Clin Invest. 1993;91(4):1769-1774. (PubMed)
65. Vergnaud P, Garnero P, Meunier PJ, Breart G, Kamihagi K, Delmas PD. Undercarboxylated osteocalcin measured with a specific immunoassay predicts hip fracture in elderly women: the EPIDOS Study. J Clin Endocrinol Metab. 1997;82(3):719-724. (PubMed)
66. Shearer MJ. The roles of vitamins D and K in bone health and osteoporosis prevention. Proc Nutr Soc. 1997;56(3):915-937. (PubMed)
67. O'Connor E, Molgaard C, Michaelsen KF, Jakobsen J, Cashman KD. Vitamin D-vitamin K interaction: effect of vitamin D supplementation on serum percentage undercarboxylated osteocalcin, a sensitive measure of vitamin K status, in Danish girls. Br J Nutr. 2010;104(8):1091-1095. (PubMed)
68. Kanellakis S, Moschonis G, Tenta R, et al. Changes in parameters of bone metabolism in postmenopausal women following a 12-month intervention period using dairy products enriched with calcium, vitamin D, and phylloquinone (vitamin K(1)) or menaquinone-7 (vitamin K (2)): the Postmenopausal Health Study II. Calcif Tissue Int. 2012;90(4):251-262. (PubMed)
69. Bolton-Smith C, McMurdo ME, Paterson CR, et al. Two-year randomized controlled trial of vitamin K1 (phylloquinone) and vitamin D3 plus calcium on the bone health of older women. J Bone Miner Res. 2007;22(4):509-519. (PubMed)
70. Braam LA, Knapen MH, Geusens P, et al. Vitamin K1 supplementation retards bone loss in postmenopausal women between 50 and 60 years of age. Calcif Tissue Int. 2003;73(1):21-26. (PubMed)
71. Orimo H, Nakamura T, Hosoi T, et al. Japanese 2011 guidelines for prevention and treatment of osteoporosis--executive summary. Arch Osteoporos. 2012;7:3-20. (PubMed)
72. Ronn SH, Harslof T, Oei L, Pedersen SB, Langdahl BL. The effect of vitamin MK-7 on bone mineral density and microarchitecture in postmenopausal women with osteopenia, a 3-year randomized, placebo-controlled clinical trial. Osteoporos Int. 2021;32(1):185-191. (PubMed)
73. Mott A, Bradley T, Wright K, et al. Effect of vitamin K on bone mineral density and fractures in adults: an updated systematic review and meta-analysis of randomised controlled trials. Osteoporos Int. 2019;30(8):1543-1559. (PubMed)
74. Jamal SA, Browner WS, Bauer DC, Cummings SR. Warfarin use and risk for osteoporosis in elderly women. Study of Osteoporotic Fractures Research Group. Ann Intern Med. 1998;128(10):829-832. (PubMed)
75. Caraballo PJ, Heit JA, Atkinson EJ, et al. Long-term use of oral anticoagulants and the risk of fracture. Arch Intern Med. 1999;159(15):1750-1756. (PubMed)
76. Gage BF, Birman-Deych E, Radford MJ, Nilasena DS, Binder EF. Risk of osteoporotic fracture in elderly patients taking warfarin: results from the National Registry of Atrial Fibrillation 2. Arch Intern Med. 2006;166(2):241-246. (PubMed)
77. Caraballo PJ, Gabriel SE, Castro MR, Atkinson EJ, Melton LJ, 3rd. Changes in bone density after exposure to oral anticoagulants: a meta-analysis. Osteoporos Int. 1999;9(5):441-448. (PubMed)
78. Fusaro M, Crepaldi G, Maggi S, et al. Bleeding, vertebral fractures and vascular calcifications in patients treated with warfarin: hope for lower risks with alternative therapies. Curr Vasc Pharmacol. 2011;9(6):763-769. (PubMed)
79. Cheung CL, Sahni S, Cheung BM, Sing CW, Wong IC. Vitamin K intake and mortality in people with chronic kidney disease from NHANES III. Clin Nutr. 2015;34(2):235-240. (PubMed)
80. Juanola-Falgarona M, Salas-Salvado J, Martinez-Gonzalez MA, et al. Dietary intake of vitamin K is inversely associated with mortality risk. J Nutr. 2014;144(5):743-750. (PubMed)
81. Zwakenberg SR, den Braver NR, Engelen AIP, et al. Vitamin K intake and all-cause and cause specific mortality. Clin Nutr. 2017;36(5):1294-1300. (PubMed)
82. Palmer CR, Bellinge JW, Dalgaard F, et al. Association between vitamin K1 intake and mortality in the Danish Diet, Cancer, and Health cohort. Eur J Epidemiol. 2021;36(10):1005-1014. (PubMed)
83. Shea MK, Barger K, Booth SL, et al. Vitamin K status, cardiovascular disease, and all-cause mortality: a participant-level meta-analysis of 3 US cohorts. Am J Clin Nutr. 2020;111(6):1170-1177. (PubMed)
84. Rees K, Guraewal S, Wong YL, et al. Is vitamin K consumption associated with cardio-metabolic disorders? A systematic review. Maturitas. 2010;67(2):121-128. (PubMed)
85. Gast GC, de Roos NM, Sluijs I, et al. A high menaquinone intake reduces the incidence of coronary heart disease. Nutr Metab Cardiovasc Dis. 2009;19(7):504-510. (PubMed)
86. Geleijnse JM, Vermeer C, Grobbee DE, et al. Dietary intake of menaquinone is associated with a reduced risk of coronary heart disease: the Rotterdam Study. J Nutr. 2004;134(11):3100-3105. (PubMed)
87. Haugsgjerd TR, Egeland GM, Nygard OK, et al. Association of dietary vitamin K and risk of coronary heart disease in middle-age adults: the Hordaland Health Study Cohort. BMJ Open. 2020;10(5):e035953. (PubMed)
88. Bellinge JW, Dalgaard F, Murray K, et al. Vitamin K intake and atherosclerotic cardiovascular disease in the Danish Diet Cancer and Health Study. J Am Heart Assoc. 2021;10(16):e020551. (PubMed)
89. Otsuka F, Sakakura K, Yahagi K, Joner M, Virmani R. Has our understanding of calcification in human coronary atherosclerosis progressed? Arterioscler Thromb Vasc Biol. 2014;34(4):724-736. (PubMed)
90. Chen J, Budoff MJ, Reilly MP, et al. Coronary artery calcification and risk of cardiovascular disease and death among patients with chronic kidney disease. JAMA Cardiol. 2017;2(6):635-643. (PubMed)
91. Rennenberg RJ, Kessels AG, Schurgers LJ, van Engelshoven JM, de Leeuw PW, Kroon AA. Vascular calcifications as a marker of increased cardiovascular risk: a meta-analysis. Vasc Health Risk Manag. 2009;5(1):185-197. (PubMed)
92. Jie KS, Bots ML, Vermeer C, Witteman JC, Grobbee DE. Vitamin K intake and osteocalcin levels in women with and without aortic atherosclerosis: a population-based study. Atherosclerosis. 1995;116(1):117-123. (PubMed)
93. Villines TC, Hatzigeorgiou C, Feuerstein IM, O'Malley PG, Taylor AJ. Vitamin K1 intake and coronary calcification. Coron Artery Dis. 2005;16(3):199-203. (PubMed)
94. Maas AH, van der Schouw YT, Beijerinck D, et al. Vitamin K intake and calcifications in breast arteries. Maturitas. 2007;56(3):273-279. (PubMed)
95. Beulens JW, Bots ML, Atsma F, et al. High dietary menaquinone intake is associated with reduced coronary calcification. Atherosclerosis. 2009;203(2):489-493. (PubMed)
96. Qiu C, Zheng H, Tao H, et al. Vitamin K2 inhibits rat vascular smooth muscle cell calcification by restoring the Gas6/Axl/Akt anti-apoptotic pathway. Mol Cell Biochem. 2017;433(1-2):149-159. (PubMed)
97. Holden RM, Hetu MF, Li TY, et al. Circulating Gas6 is associated with reduced human carotid atherosclerotic plaque burden in high risk cardiac patients. Clin Biochem. 2019;64:6-11. (PubMed)
98. Viegas CS, Rafael MS, Enriquez JL, et al. Gla-rich protein acts as a calcification inhibitor in the human cardiovascular system. Arterioscler Thromb Vasc Biol. 2015;35(2):399-408. (PubMed)
99. Borras T, Comes N. Evidence for a calcification process in the trabecular meshwork. Exp Eye Res. 2009;88(4):738-746. (PubMed)
100. Schurgers LJ, Uitto J, Reutelingsperger CP. Vitamin K-dependent carboxylation of matrix Gla-protein: a crucial switch to control ectopic mineralization. Trends Mol Med. 2013;19(4):217-226. (PubMed)
101. Cassidy-Bushrow AE, Bielak LF, Levin AM, et al. Matrix gla protein gene polymorphism is associated with increased coronary artery calcification progression. Arterioscler Thromb Vasc Biol. 2013;33(3):645-651. (PubMed)
102. Herrmann SM, Whatling C, Brand E, et al. Polymorphisms of the human matrix gla protein (MGP) gene, vascular calcification, and myocardial infarction. Arterioscler Thromb Vasc Biol. 2000;20(11):2386-2393. (PubMed)
103. Schurgers LJ, Teunissen KJ, Knapen MH, et al. Novel conformation-specific antibodies against matrix gamma-carboxyglutamic acid (Gla) protein: undercarboxylated matrix Gla protein as marker for vascular calcification. Arterioscler Thromb Vasc Biol. 2005;25(8):1629-1633. (PubMed)
104. van den Heuvel EG, van Schoor NM, Lips P, et al. Circulating uncarboxylated matrix Gla protein, a marker of vitamin K status, as a risk factor of cardiovascular disease. Maturitas. 2014;77(2):137-141. (PubMed)
105. Mayer O, Jr., Seidlerova J, Bruthans J, et al. Desphospho-uncarboxylated matrix Gla-protein is associated with mortality risk in patients with chronic stable vascular disease. Atherosclerosis. 2014;235(1):162-168. (PubMed)
106. Shea MK, O'Donnell CJ, Hoffmann U, et al. Vitamin K supplementation and progression of coronary artery calcium in older men and women. Am J Clin Nutr. 2009;89(6):1799-1807. (PubMed)
107. Shea MK, O'Donnell CJ, Vermeer C, et al. Circulating uncarboxylated matrix gla protein is associated with vitamin K nutritional status, but not coronary artery calcium, in older adults. J Nutr. 2011;141(8):1529-1534. (PubMed)
108. Ikari Y, Torii S, Shioi A, Okano T. Impact of menaquinone-4 supplementation on coronary artery calcification and arterial stiffness: an open label single arm study. Nutr J. 2016;15(1):53. (PubMed)
109. Diederichsen ACP, Lindholt JS, Moller S, et al. Vitamin K2 and D in patients with aortic valve calcification: a randomized double-blinded clinical trial. Circulation. 2022;145(18):1387-1397. (PubMed)
110. Zwakenberg SR, de Jong PA, Bartstra JW, et al. The effect of menaquinone-7 supplementation on vascular calcification in patients with diabetes: a randomized, double-blind, placebo-controlled trial. Am J Clin Nutr. 2019;110(4):883-890. (PubMed)
111. Kurnatowska I, Grzelak P, Masajtis-Zagajewska A, et al. Effect of vitamin K2 on progression of atherosclerosis and vascular calcification in nondialyzed patients with chronic kidney disease stages 3-5. Pol Arch Med Wewn. 2015;125(9):631-640. (PubMed)
112. De Vriese AS, Caluwe R, Pyfferoen L, et al. Multicenter randomized controlled trial of vitamin K antagonist replacement by rivaroxaban with or without vitamin K2 in hemodialysis patients with atrial fibrillation: the Valkyrie Study. J Am Soc Nephrol. 2020;31(1):186-196. (PubMed)
113. Lees JS, Chapman FA, Witham MD, Jardine AG, Mark PB. Vitamin K status, supplementation and vascular disease: a systematic review and meta-analysis. Heart. 2019;105(12):938-945. (PubMed)
114. Chatrou ML, Winckers K, Hackeng TM, Reutelingsperger CP, Schurgers LJ. Vascular calcification: the price to pay for anticoagulation therapy with vitamin K-antagonists. Blood Rev. 2012;26(4):155-166. (PubMed)
115. US Centers for Disease Control and Prevention, National Center for Chronic Disease Prevention and Health Promotion, Division of Population Health,. Osteoarthritis (OA). Available at: https://www.cdc.gov/arthritis/basics/osteoarthritis.htm. Accessed 5/19/22.
116. Harshman SG, Shea MK. The role of vitamin K in chronic aging diseases: inflammation, cardiovascular disease, and osteoarthritis. Curr Nutr Rep. 2016;5(2):90-98. (PubMed)
117. Oka H, Akune T, Muraki S, et al. Association of low dietary vitamin K intake with radiographic knee osteoarthritis in the Japanese elderly population: dietary survey in a population-based cohort of the ROAD study. J Orthop Sci. 2009;14(6):687-692. (PubMed)
118. Neogi T, Booth SL, Zhang YQ, et al. Low vitamin K status is associated with osteoarthritis in the hand and knee. Arthritis Rheum. 2006;54(4):1255-1261. (PubMed)
119. Misra D, Booth SL, Tolstykh I, et al. Vitamin K deficiency is associated with incident knee osteoarthritis. Am J Med. 2013;126(3):243-248. (PubMed)
120. Shea MK, Kritchevsky SB, Hsu FC, et al. The association between vitamin K status and knee osteoarthritis features in older adults: the Health, Aging and Body Composition Study. Osteoarthritis Cartilage. 2015;23(3):370-378. (PubMed)
121. Boer CG, Szilagyi I, Nguyen NL, et al. Vitamin K antagonist anticoagulant usage is associated with increased incidence and progression of osteoarthritis. Ann Rheum Dis. 2021;80(5):598-604. (PubMed)
122. Neogi T, Felson DT, Sarno R, Booth SL. Vitamin K in hand osteoarthritis: results from a randomised clinical trial. Ann Rheum Dis. 2008;67(11):1570-1573. (PubMed)
123. Harshman SG, Finnan EG, Barger KJ, et al. Vegetables and mixed dishes are top contributors to phylloquinone intake in US adults: data from the 2011-2012 NHANES. J Nutr. 2017;147(7):1308-1313. (PubMed)
124. Vermeer C, Raes J, van 't Hoofd C, Knapen MHJ, Xanthoulea S. Menaquinone content of cheese. Nutrients. 2018;10(4). (PubMed)
125. Walther B, Karl JP, Booth SL, Boyaval P. Menaquinones, bacteria, and the food supply: the relevance of dairy and fermented food products to vitamin K requirements. Adv Nutr. 2013;4(4):463-473. (PubMed)
126. Ellis JL, Fu X, Karl JP, et al. Multiple dietary vitamin K forms are converted to tissue menaquinone-4 in mice. J Nutr. 2022;152(4):981-993. (PubMed)
127. Beulens JW, Booth SL, van den Heuvel EG, Stoecklin E, Baka A, Vermeer C. The role of menaquinones (vitamin K(2)) in human health. Br J Nutr. 2013;110(8):1357-1368. (PubMed)
128. US Department of Health and Human Services, National Institutes of Health, Office of Dietary Supplements. Dietary Supplement Label Database (DSLD). [Internet]. Accessed 3/7/22. Available from: https://dsld.od.nih.gov/.
129. Traber MG. Vitamin E and K interactions--a 50-year-old problem. Nutr Rev. 2008;66(11):624-629. (PubMed)
130. Booth SL, Golly I, Sacheck JM, et al. Effect of vitamin E supplementation on vitamin K status in adults with normal coagulation status. Am J Clin Nutr. 2004;80(1):143-148. (PubMed)
131. Pastori D, Carnevale R, Cangemi R, et al. Vitamin E serum levels and bleeding risk in patients receiving oral anticoagulant therapy: a retrospective cohort study. J Am Heart Assoc. 2013;2(6):e000364. (PubMed)
132. Shearer MJ, Newman P. Recent trends in the metabolism and cell biology of vitamin K with special reference to vitamin K cycling and MK-4 biosynthesis. J Lipid Res. 2014;55(3):345-362. (PubMed)
133. Chang CH, Wang YW, Yeh Liu PY, Kao Yang YH. A practical approach to minimize the interaction of dietary vitamin K with warfarin. J Clin Pharm Ther. 2014;39(1):56-60. (PubMed)
134. Thorp JA, Gaston L, Caspers DR, Pal ML. Current concepts and controversies in the use of vitamin K. Drugs. 1995;49(3):376-387. (PubMed)
135. Reiffel JA. An important indirect drug interaction between dronedarone and warfarin that may be extrapolated to other drugs that can alter gastrointestinal function. Am Heart J. 2011;161(2):e5; author reply e7. (PubMed)
136. Shirolkar SC, Fiuzat M, Becker RC. Dronedarone and vitamin K antagonists: a review of drug-drug interactions. Am Heart J. 2010;160(4):577-582. (PubMed)
137. Hendler SS, Rorvik DR, eds. PDR for Nutritional Supplements. Montvale: Medical Economics Company, Inc.; 2001.
If you appreciate this website, please help us maintain and update the Micronutrient Information Center. Make a gift during Oregon State University's annual giving drive.