Resumen
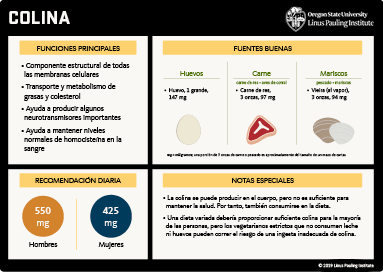
- La colina es un nutriente esencial similar a una vitamina y un donante de metilo involucrado en muchos procesos fisiológicos, incluido el metabolismo normal y el transporte de lípidos, reacciones de metilación y síntesis de neurotransmisores. (Más información)
- La deficiencia de colina causa un depósito anormal de grasa en el hígado, lo que resulta en una condición llamada enfermedad del hígado graso no alcohólico. En algunas personas, la deficiencia de colina provoca daño muscular. Las variantes genéticas, el sexo y el estado hormonal influyen en las necesidades individuales y, por tanto, en la susceptibilidad a la enfermedad del hígado graso inducida por la deficiencia de colina. (Más información)
- La ingesta adecuada (IA) recomendada de colina para estadounidenses y canadienses es diferente según los grupos de edad y el sexo. Para los adultos, la ingesta recomendada es de 425 miligramos (mg)/día para las mujeres y 550 mg/día para los hombres. Las recomendaciones de ingesta dietética aumentan durante el embarazo a 450 mg/día y durante la lactancia a 550 mg/día. (Más información) La gran mayoría de las personas que viven en los Estados Unidos tienen ingestas dietéticas inferiores al IA. (Más información)
- La colina participa en la regulación de la concentración de homocisteína en la sangre a través de su metabolito betaína. Actualmente no hay evidencia convincente de que el consumo elevado de colina pueda beneficiar la salud cardiovascular al reducir la homocisteína en la sangre. (Más información) Además, las concentraciones sanguíneas elevadas de N-óxido de trimetilamina (TMAO), generado a partir de colina, se han asociado con un mayor riesgo de eventos cardiovasculares en algunos estudios observacionales. (Más información)
- La necesidad de colina probablemente aumenta durante el embarazo. Los estudios de caso y control que examinan la relación entre el estado materno de colina y el riesgo de defectos del tubo neural han arrojado resultados inconsistentes. Aún no se sabe si la suplementación con colina durante el período periconcepcional podría conferir protección contra los defectos del tubo neural. (Más información)
- Los estudios en animales han demostrado que la colina es esencial para el desarrollo óptimo del cerebro e influye en la función cognitiva en la vejez. Sin embargo, se necesitan más estudios en humanos para afirmar que la suplementación con colina durante el embarazo mejora el rendimiento cognitivo de la descendencia o que la suplementación con colina ayuda a prevenir el deterioro cognitivo en las personas mayores. (Más información)
- Si bien los resultados de los estudios de intervención son contradictorios, algunas pruebas sugieren que el tratamiento con citicolina (un derivado de la colina) puede ser útil para mejorar la función de la retina en algunos pacientes con glaucoma. La citicolina también se ha estudiado como un tratamiento potencial para limitar el daño neurológico en pacientes que sufren un accidente cerebrovascular o una lesión cerebral traumática. (Más información)
- La síntesis de novo de colina en humanos no es suficiente para satisfacer las necesidades metabólicas; por lo tanto, es necesaria la ingesta dietética de colina. Buenas fuentes dietéticas de colina incluyen huevos, carne, aves, pescado, vegetales crucíferos, maní y productos lácteos. (Más información)
- El consumo excesivo de colina (≥7,500 mg) se ha asociado con la disminución de la presión arterial, sudoración, olor corporal a pescado, y efectos secundarios gastrointestinales. El nivel máximo de ingesta tolerable (NM) para adultos es de 3,500 mg/día. (Más información)
Aunque la colina no es, por definición estricta una vitamina, es un nutriente esencial. A pesar de que los humanos podemos sintetizarla en pequeñas cantidades, la colina debe consumirse en la dieta para mantener una buena salud. La mayoría de la colina del cuerpo se encuentra en moléculas de grasa especializadas conocidas como fosfolípidos, de los cuales la más común se llama fosfatidilcolina (1).
Función
La colina y los compuestos derivados de la colina (p. ej., los metabolitos) cumplen una serie de funciones biológicas vitales (Figura 1) (1).

Integridad estructural de las membranas celulares
La colina se utiliza en la síntesis de ciertos fosfolípidos (fosfatidilcolina y esfingomielina) que son componentes estructurales esenciales de las membranas celulares. La fosfatidilcolina representa aproximadamente el 95% de la colina total en los tejidos (2). Este fosfolípido se puede sintetizar a partir de la colina dietética mediante la vía de la citidina difosfocolina (CDP-colina) o mediante la metilación de otro fosfolípido, la fosfatidiletanolamina (Figura 2) (3). La esfingomielina es un tipo de fosfolípido que contiene esfingosina (esfingolípido) que se sintetiza mediante la transferencia de un residuo de fosfocolina de la fosfatidilcolina a una ceramida (Figura 3). La esfingomielina se encuentra en las membranas celulares y en la vaina grasa que envuelve las fibras nerviosas mielinizadas.



Señalización celular
Los fosfolípidos que contienen colina, la fosfatidilcolina y la esfingomielina, son precursores de las moléculas mensajeras intracelulares, el diacilglicerol y la ceramida. Específicamente, las esfingomielinasas (también conocidas como esfingomielina fosfodiesterasas) catalizan la escisión de la esfingomielina, generando fosfocolina y ceramida. El diacilglicerol se libera mediante la degradación de la fosfatidilcolina por las fosfolipasas. Otros metabolitos de la colina que se sabe que son moléculas de señalización celular incluyen el factor activador de plaquetas (PAF) y la esfingofosfocolina.
Transmisión de impulso nervioso
La colina es un precursor de la acetilcolina, un importante neurotransmisor sintetizado por neuronas colinérgicas e implicado en el control muscular, el ritmo circadiano, la memoria, y muchas otras funciones neuronales. La colina acetiltransferasa cataliza la acetilación de colina a acetilcolina, y la acetilcolina esterasa hidroliza la acetilcolina a colina y acetato (4). También se descubrió que la administración de CDP-colina (citicolina) estimula la síntesis y liberación de una familia de neurotransmisores derivados de la tirosina (es decir, las catecolaminas, incluidas noradrenalina, adrenalina y dopamina) (5). Es de destacar que las células no neuronales de diversos tejidos y sistemas de órganos también sintetizan y liberan acetilcolina, que luego se une y estimula los receptores colinérgicos en las células diana (revisado en 6).
Transporte y metabolismo de lípidos (grasas)
La grasa y el colesterol consumidos en la dieta son transportados al hígado mediante lipoproteínas llamadas quilomicrones. En el hígado, la grasa y el colesterol se empaquetan en lipoproteínas llamadas lipoproteínas de muy baja densidad (VLDL) para su transporte en el torrente sanguíneo a los tejidos extrahepáticos. La síntesis de fosfatidilcolina por la vía de la fosfatidiletanolamina N-metiltransferasa (PEMT) es necesaria para el ensamblaje y la secreción de VLDL en el hígado (7, 8). Los polimorfismos del gen PEMT aumentan los requerimientos dietéticos de colina (9). Sin una fosfatidilcolina adecuada, la grasa y el colesterol se acumulan en el hígado (ver Deficiencia).
Fuente principal de grupos metilo
La colina puede oxidarse en el hígado y los riñones para formar un metabolito llamado betaína mediante una reacción enzimática de dos pasos. En la membrana interna de la mitocondria, la colina oxidasa dependiente de dinucleótido de flavina adenina (FAD) cataliza la conversión de colina en betaína aldehído, que luego es convertida en betaína por la betaína aldehído deshidrogenasa en la matriz mitocondrial o en el citosol (2). La betaína es una fuente de hasta el 60% de los grupos metilo (CH3) necesarios para la metilación de la homocisteína (10). La betaína homocisteína metiltransferasa (BHMT) utiliza la betaína como donante de metilo para convertir la homocisteína en metionina en el metabolismo de un carbono (Figura 4). La ubicua enzima metionina sintasa (MS) dependiente de la vitamina B12 también cataliza la remetilación de la homocisteína, utilizando el derivado de folato, 5-metiltetrahidrofolato, como donante de metilo (ver Interacciones de nutrientes). Las concentraciones elevadas de homocisteína en la sangre se han asociado con un mayor riesgo de enfermedad cardiovascular (11).

Osmorregulación
La conversión de colina en betaína es irreversible. La betaína es un osmolito que regula el volumen celular y protege la integridad celular contra el estrés osmótico (especialmente en el riñón). El estrés osmótico se ha asociado con una expresión reducida de BHMT, de modo que el papel de la betaína en la osmorregulación puede priorizarse temporalmente sobre su función como donante de metilo (2).
Deficiencia
Síntomas
Se ha descubierto que hombres y mujeres alimentados por vía intravenosa (IV) con soluciones que contenían metionina y folato adecuados pero que carecían de colina desarrollaron una condición llamada enfermedad del hígado graso no alcohólico (NAFLD) y signos de daño hepático que se resolvieron cuando se suministró colina (12). La aparición de La NAFLD generalmente se asocia con la presentación conjunta de trastornos metabólicos, incluida la obesidad, la dislipidemia, la resistencia a la insulina, y la hipertensión, en sujetos con síndrome metabólico. Se estima que la NAFLD progresa a una afección más grave llamada esteatohepatitis no alcohólica (NASH) en aproximadamente un tercio de los pacientes con NAFLD, además de aumentar el riesgo de cirrosis y cáncer de hígado (13).
Debido a que la fosfatidilcolina es necesaria para la síntesis de partículas de lipoproteínas de muy baja densidad (VLDL) (ver Función), la deficiencia de colina produce una secreción alterada de VLDL y acumulación de grasa en el hígado (esteatosis), lo que en última instancia conduce a daño hepático. Debido a que las partículas de lipoproteínas de baja densidad (LDL) se forman a partir de partículas de VLDL, las personas con deficiencia de colina también muestran concentraciones reducidas de colesterol LDL en la sangre (14). Los biomarcadores anormalmente elevados de disfunción orgánica en la sangre, incluida la creatina fosfoquinasa, la aspartato aminotransferasa y la alanina aminotransferasa, se corrigen tras la reposición de colina. La disfunción orgánica inducida por deficiencia de colina también se ha asociado con un aumento del daño en el ADN y la apoptosis en los linfocitos circulantes (15). En el hígado, se cree que la acumulación de lípidos deteriora la función mitocondrial, reduciendo así la oxidación de los ácidos grasos y aumentando la producción de especies reactivas de oxígeno (ERO) que desencadenan la peroxidación de lípidos, el daño del ADN, y la apoptosis. Además, se cree que el estrés oxidativo es responsable de provocar procesos inflamatorios que pueden conducir a la progresión de NAFLD a NASH y cirrosis (enfermedad hepática terminal) (16).
Un estudio de intervención en 57 adultos sanos que fueron alimentados con dietas deficientes en colina en condiciones controladas encontró que el 77% de los hombres, el 80% de las mujeres posmenopáusicas, y el 44% de las mujeres premenopáusicas desarrollaron hígado graso, daño hepático, y/o daño muscular (17). Estos signos de disfunción orgánica se resolvieron tras la reintroducción de colina en la dieta. Debido a que el estrógeno estimula la síntesis endógena de fosfatidilcolina a través de la vía de la fosfatidiletanolamina N-metiltransferasa (PEMT), las mujeres premenopáusicas pueden ser menos propensas a desarrollar signos de deficiencia de colina en respuesta a una dieta baja en colina en comparación con las mujeres postmenopáusicas (18, 19). Además, se cree que un notable polimorfismo de un solo nucleótido (SNP; rs12325817) del gen PEMT, que puede afectar la expresión y/o actividad de la enzima PEMT, aumenta la susceptibilidad a la disfunción orgánica inducida por deficiencia de colina (18). Los polimorfismos genéticos adicionales que ocurren en las rutas metabólicas de colina y de un carbono pueden alterar el requerimiento dietético de colina y, por lo tanto, aumentar la probabilidad de desarrollar signos de deficiencia cuando la ingesta de colina es insuficiente (9, 20, 21).
Es de destacar que la composición de la microbiota intestinal se ha identificado como un predictor potencial de susceptibilidad a la NAFLD inducida por deficiencia de colina (22). El metabolismo de la fosfatidilcolina dietética dependiente de la microbiota intestinal también podría estar involucrado en la patogénesis de la enfermedad cardiovascular (ver Seguridad) (23, 24).
Para obtener más información sobre las enfermedades del hígado graso, consulte la sección sobre Prevención de Enfermedad.
Individuos en riesgo de insuficiencia de colina
Las recomendaciones de ingesta diaria son más altas durante el embarazo y la lactancia (ver La IA), lo que coloca a las personas embarazadas y lactantes en mayor riesgo de insuficiencia de colina (25, 26). Los vegetarianos también pueden tener un mayor riesgo de insuficiencia de colina (26).
Los pacientes con fibrosis quística que tienen insuficiencia pancreática corren el riesgo de sufrir una depleción de colina debido al aumento de las pérdidas fecales (27). La colina circulante se ha correlacionado directamente con la función pulmonar en pacientes con fibrosis quística (28), y los resultados de un estudio piloto en 10 pacientes sugirieron que la suplementación con colina puede tienen utilidad para mejorar los resultados clínicos en pacientes con fibrosis quística (29).
Interacciones de nutrientes
Junto con varias vitaminas B (es decir, folato, vitamina B12, vitamina B6, y riboflavina), la colina es necesaria para el metabolismo de los ácidos nucleicos y aminoácidos, y para la generación del donante universal del grupo metilo, la S-adenosilmetionina (SAM) (ver Figura 4). SAM se sintetiza a partir del aminoácido esencial, la metionina. Se requieren tres moléculas de SAM para la reacción de metilación que convierte la fosfatidiletanolamina en fosfatidilcolina (ver Figura 2). Una vez que SAM dona un grupo metilo, se convierte en S-adenosilhomocisteína (SAH), que luego se metaboliza a homocisteína. La homocisteína se puede convertir nuevamente en metionina en una reacción catalizada por la metionina sintasa dependiente de vitamina B12, que requiere 5-metiltetrahidrofolato (5-meTHF) como donante de metilo. Alternativamente, la betaína (un metabolito de la colina) se utiliza como donante de metilo para la metilación de la homocisteína en metionina mediante la enzima, betaína-homocisteína metiltransferasa (BHMT) (1). La homocisteína también se puede metabolizar a cisteína a través de la vía de transulfuración dependiente de la vitamina B6 (ver Figura 4).
Por lo tanto, el requerimiento humano de colina está especialmente influenciado por la relación entre la colina y otros donantes de grupos metilo, como el folato y la S-adenosilmetionina. Una baja ingesta de folato conduce a una mayor demanda de de metabolito derivado de colina, la betaína. Además, la síntesis de novo de fosfatidilcolina no es suficiente para mantener un estado nutricional adecuado de colina cuando las ingestas dietéticas de folato y colina son bajas (30). Por el contrario, la demanda de folato aumenta cuando el suministro dietético de colina es limitado (31).
La IA
En 1998, la Junta de Alimentos y Nutrición (JNA) del Instituto de Medicina estableció una ingesta diaria recomendada (IDR) para la colina (32). La JNA sintió que la evidencia científica existente era insuficiente para calcular una IDR de colina, por lo que establecieron una ingesta adecuada (IA; Tabla 1). El criterio principal para establecer la IA para la colina fue la prevención del daño hepático. Sin embargo, los polimorfismos comunes en los genes involucrados en el metabolismo de la colina o el folato alteran la susceptibilidad a la deficiencia de colina y, por lo tanto, pueden afectar los requerimientos dietarios de colina (ver Deficiencia) (18, 20, 33, 34).
| Grupo Etario | Edad | Machos (mg/día) | Hembras (mg/día) |
|---|---|---|---|
| Infantes | 0-6 meses | 125 | 125 |
| Infantes | 7-12 meses | 150 | 150 |
| Niños | 1-3 años | 200 | 200 |
| Niños | 4-8 años | 250 | 250 |
| Niños | 9-13 años | 375 | 375 |
| Adolescentes | 14-18 años | 550 | 400 |
| Adultos | 19 años y más | 550 | 425 |
| Embarazo | Todas las edades | - | 450 |
| Período de lactancia | Todas las edades | - | 550 |
Prevención de Enfermedad
Enfermedad cardiovascular
Colina y homocisteína
Una gran cantidad de investigación indica que incluso niveles moderadamente elevados de homocisteína en la sangre aumentan el riesgo de enfermedad cardiovascular (11). La causa más común de infarto al miocardio o accidente cerebrovascular es la ruptura de las placas ateroscleróticas en las paredes arteriales que causan la formación de coágulos sanguíneos (trombogénesis). Las altas concentraciones de homocisteína pueden promover el desarrollo de aterosclerosis (aterogénesis) y trombogénesis a través de mecanismos que involucran estrés oxidativo y disfunción endotelial, inflamación, coagulación sanguínea anormal, y metabolismo lipídico desordenado (revisado en 35).
Una vez formada a partir de metionina dietética, la homocisteína se puede catabolizar a cisteína a través de la vía de transulfuración o remetilarse en metionina (ver Figura 4). El folato y la colina están involucrados en rutas alternativas que catalizan la remetilación de la homocisteína (ver Interacciones de nutrientes). De manera específica, la colina es el precursor de la betaína, que proporciona un grupo metilo para la conversión de homocisteína en metionina a través de la enzima, betaína-homocisteína metiltransferasa (BHMT). Si bien la cantidad de homocisteína en la sangre está regulada por varios nutrientes, incluidos el folato y la colina, las condiciones que causan daño al hígado como la esteatohepatitis no alcohólica (EHNA) también pueden afectar el metabolismo de la homocisteína (36).
Ingestas dietéticas de colina y betaína y ECV
Debido a que las rutas metabólicas dependientes de folato y colina catalizan la remetilación de la homocisteína, es necesario considerar la ingesta dietética de ambos nutrientes cuando se evalúa la asociación entre las concentraciones de homocisteína y la enfermedad cardiovascular. Sin embargo, a pesar de su relevancia, la relación de la betaína y la colina con el metabolismo de la homocisteína solo se ha investigado ligeramente en humanos, esencialmente porque el contenido de colina de los alimentos no se podía medir con precisión hasta hace poco. En estudios de intervención preliminares, se encontró que las dosis farmacológicas de betaína (1,500 a 6,000 mg/día) redujeron las concentraciones de homocisteína en la sangre en un pequeño número de voluntarios con concentraciones de homocisteína normales a moderadamente elevadas (37-40). Sin embargo, en un análisis de corte transversal de una gran cohorte de 16,165 mujeres (edades, 49-79 años), no se encontró que dosis más bajas de betaína en el rango de ingestas dietéticas estuvieran correlacionadas con las concentraciones de homocisteína (41). Este estudio también mostró que los niveles de ingesta de colina estaban inversamente asociados con las concentraciones de homocisteína en la sangre. Sin embargo, un estudio de seguimiento de ocho años de la cohorte no mostró ninguna diferencia en el riesgo de enfermedad cardiovascular entre las mujeres en el cuartil superior versus inferior de la ingesta de colina dietética (>329 mg/día frente a ≤266 mg/día) (41).
Los estudios prospectivos de cohorte más recientes sobre la asociación de la colina o betaína dietética y las enfermedades cardiovasculares han sido mixtos. En el Estudio de Jackson Heart, que siguió a 3,924 afroamericanos durante nueve años, una mayor ingesta de colina en la dieta se asoció con un riesgo reducido de accidente cerebrovascular (42). En el Estudio de Salud de Enfermeras (Nurses’ Health Study) (80, 978 mujeres) y el Estudio de Seguimiento de Profesionales de la Salud (Health Professionals Follow-up Study) (39,434 hombres), una mayor ingesta dietética de fosfatidilcolina se relacionó con un mayor riesgo de mortalidad relacionada con enfermedades cardiovasculares (43). Una mayor ingesta diaria total de colina se relacionó con un mayor riesgo de mortalidad cardiometabólica (es decir, muerte atribuida a enfermedades cardiovasculares o diabetes mellitus tipo 2) en grandes cohortes prospectivas de tres grupos étnicos: negros (n=49,858), blancos (n=23,766), y chinos (n=134,001) (44). En este estudio, una mayor ingesta de betaína se asoció con un mayor riesgo de muerte cardiometabólica solo en la cohorte china (44). Sin embargo, no se encontraron asociaciones entre la ingesta de colina o betaína y la mortalidad relacionada con enfermedades cardiovasculares en un estudio de cohorte de adultos japoneses (n=29,079) (45). Varios otros estudios de cohortes prospectivos no han logrado encontrar una asociación entre la ingesta de colina y los criterios de evaluación de enfermedades cardiovasculares (41, 46-48).
Actualmente falta evidencia convincente de que una mayor ingesta dietética de colina o betaína podría beneficiar la salud cardiovascular al reducir las concentraciones de homocisteína en la sangre.
Concentraciones circulantes de colina y betaína y riesgo de enfermedad cardiovascular
Un estudio de 1995 encontró que las concentraciones elevadas de homocisteína en sangre en pacientes que experimentaron una oclusión vascular se asociaron con una mayor excreción urinaria de betaína, en vez de con una ingesta reducida de colina o betaína o una actividad disminuida de BHMT (49). En un estudio prospectivo reciente, un alto nivel de excreción urinaria de betaína también se asoció con un mayor riesgo de insuficiencia cardíaca en 325 sujetos sin diabetes mellitus que fueron hospitalizados por síndrome coronario agudo (50). En el mismo estudio, los quintiles superior e inferior de las concentraciones en plasma de betaína se asociaron con un mayor riesgo de infarto al miocardio agudo secundario. Los hallazgos de otro estudio prospectivo (el Estudio de Salud Hordaland; Hordaland Health Study) que siguió a 7,045 adultos sanos (edades, 47-49 años y 71-74 años) sugirieron que las concentraciones altas de colina y betaína en plasma se asociaron con un perfil desfavorable de riesgo cardiovascular (51). De hecho , la colina en plasma se asoció positivamente con una serie de factores de riesgo cardiovascular, como el IMC, el porcentaje de grasa corporal, la circunferencia de la cintura, y los triglicéridos séricos, e inversamente asociado con el colesterol HDL. Por el contrario, la betaína en plasma se correlacionó positivamente con el colesterol HDL e inversamente con los factores de riesgo antes mencionados, así como con la presión arterial sistólica y diastólica.
Estudios más recientes sugieren que la concentración sanguínea de N-óxido de trimetilamina (TMAO), generada a partir de nutrientes que contienen trimetilamina como la colina dietética, en lugar de la colina, podría influir en el riesgo de eventos cardiovasculares (ver Seguridad). Sin embargo, no se observó asociación entre niveles elevados de TMAO en la sangre y aterosclerosis en el Estudio Desarrollo de Arterias Coronarias en Adulto Jóvenes (Coronary Artery Risk Development in Young Adults Study) (CARDIA) (52).
Aún no está claro si las concentraciones de colina, betaína y/o TMAO en la sangre pueden predecir el riesgo de enfermedad cardiovascular.
Enfermedades hepáticas
Enfermedades del hígado graso
Si bien una dieta deficiente en colina produce disfunción orgánica y enfermedad del hígado graso no alcohólico (NAFLD) (ver Deficiencia; 17), no se sabe si una ingesta dietética de colina subóptima en sujetos sanos puede contribuir a un mayor riesgo de NAFLD. Se realizó un análisis de corte transversal de dos grandes estudios prospectivos realizados en China — el Shanghai Women’s Health Study y el Shanghai Men’s Health Study — que incluyeron a 56,195 personas (edades, 40-75 años), para evaluar la asociación entre la ingesta dietética de colina y el diagnóstico autoinformado de enfermedad del hígado graso (53). El quintil más alto versus el más bajo de ingesta de colina (412 mg/día versus 179 mg/día) se asoció con un riesgo 28% menor de enfermedad del hígado graso en mujeres con peso normal, pero no se encontró asociación en mujeres con sobrepeso u obesidad o en hombres. Otro estudio de corte transversal de de 664 personas con NAFLD o esteatohepatitis no alcohólica (NASH) también informó que la gravedad de la enfermedad estaba inversamente correlacionada con la ingesta dietética de colina en mujeres posmenopáusicas, pero no en mujeres premenopáusicas, hombres o niños (54). Además, en una encuesta de corte transversal nacional de EE. UU. de 20,643 adultos, una mayor ingesta dietética de colina se asoció con un perfil más favorable de enzimas hepáticas y con un menor riesgo de desarrollar NAFLD (NHANES 2005-2012) (55).
Cáncer de hígado
En modelos animales, la deficiencia de colina dietética se ha asociado con una mayor incidencia de cáncer espontáneo de hígado (carcinoma hepatocelular) y una mayor sensibilidad a los productos químicos cancerígenos (10). Se han propuesto varios mecanismos para contribuir a los efectos promotores del cáncer de la deficiencia de colina: (1) mejora de la regeneración de las células hepáticas y la sensibilidad de los tejidos a las agresiones químicas; (2) expresión alterada de numerosos genes que regulan la proliferación celular, la diferenciación, la reparación del ADN, y la apoptosis debido a la metilación inadecuada del ADN; (3) mayor probabilidad de daño en el ADN causado por el estrés oxidativo inducido por disfunción mitocondrial; y (4) la cascada de señalización celular mediada por proteína quinasa C activada, que finalmente conduce a un aumento de la apoptosis de las células hepáticas (2). Sin embargo, no se sabe si la deficiencia de colina puede aumentar la susceptibilidad al cáncer en humanos (2).
Defectos del tubo neural
Se sabe que el folato es crítico para el desarrollo embrionario normal, y la suplementación materna con ácido fólico disminuye la incidencia de defectos del tubo neural (DTNs) (56). Los DTNs incluyen diversas malformaciones, como lesiones del cerebro (p. ej., anencefalia, encefalocele) o lesiones de la columna vertebral (espina bífida), que son devastadores y generalmente incompatibles con la vida (57). Estos defectos ocurren entre los días 21 y 28 después de la concepción, un momento en que muchas mujeres no se dan cuenta de que están embarazadas (58). Mientras que el efecto protector del folato contra DTN está bien establecido, solo unos pocos estudios han investigado el papel de otros donantes de grupos metilo, incluyendo colina y betaína, en la aparición de DTNs. Un estudio de caso y control (424 casos de DTN y 440 controles) encontró que las mujeres en el cuartil más alto versus el más bajo de la ingesta periconcepcional de colina (>498.46 mg/día vs. ≤290.41 mg/día) tenían un riesgo 51% menor de embarazo afectado por DTN (59). Sin embargo, estudios más recientes no lograron encontrar una relación inversa entre la ingesta materna de colina y el riesgo de DTN (60-63).
Un estudio de caso y control (80 embarazos afectados por DTN y 409 controles) en mujeres de EE.UU. encontró que las concentraciones séricas más bajas de colina (<2.49 mmol/L) durante la mitad del embarazo se asoció con un riesgo 2.4 veces mayor de DTN (64). Finalmente, un estudio más reciente, que incluye 71 embarazos afectados por DTN, 214 embarazos con malformaciones no por DTN, 98 embarazos normales en mujeres con embarazos previos afectados por DTN, y 386 embarazos normales, no encontró asociaciones entre las concentraciones de colina en la sangre materna durante el embarazo, las variantes genéticas relacionadas con colina y folato, y el riesgo de DTN (65). Sin embargo, es importante señalar que las concentraciones de colina circulante no reflejan con precisión la ingesta dietética de colina.
En un meta-análisis reciente que combinó los resultados de cinco estudios de casos y controles (59, 60, 62-64), incluyendo 1,131 embarazos afectados por DTN y 4,439 controles sanos, una menor ingesta de colina dietética o menores concentraciones séricas de colina se asociaron con un 36% de riesgo mayor de DTN en comparación con niveles más altos (95% IC, 1.11, 1.67) (66).
Se necesitarían ensayos controlados aleatorios sobre la suplementación con colina durante el período periconcepcional para determinar si la colina tiene un efecto protector contra los DTNs.
Salud cognitiva
Desarrollo neurocognitivo
El aumento de la ingesta dietética de citidina 5’-difosfocolina (CDP-colina o citicolina, un precursor de fosfatidilcolina; ver Figura 2) muy temprano en la vida puede disminuir la gravedad de los déficits de memoria en ratas de edad avanzada (67). La suplementación con colina de las madres de ratas no nacidas, así como las crías de rata durante el primer mes de vida, condujo a un mejor rendimiento en las pruebas de memoria espacial meses después de que se suspendiera la suplementación con colina (68). Una revisión de McCann et al. analiza la evidencia experimental de los estudios con roedores sobre la disponibilidad de colina durante el desarrollo prenatal y la función cognitiva en la descendencia (69).
Debido a la importancia de la metilación del ADN en el desarrollo normal del cerebro, las funciones neuronales, y los procesos cognitivos (70), los nutrientes de los donantes de metilo como la colina son esenciales para el funcionamiento óptimo del cerebro. Sin embargo, la evidencia clínica para determinar si los hallazgos en los estudios de roedores son aplicables a humanos actualmente es limitada. Recientemente, el análisis del estudio de Seychelles Child Development Nutrition Cohort reportó la falta de una asociación entre las concentraciones en plasma de colina y sus metabolitos relacionados y las capacidades cognitivas en 256 niños de cinco años. Solo se encontró que las concentraciones en plasma de betaína se correlacionan positivamente con las puntuaciones de pruebas preescolares de idioma (71). Sin embargo, debido a que las concentraciones circulantes de colina no están directamente relacionadas con la ingesta dietética de colina, el estudio no pudo evaluar si las ingestas maternas de colina influyen en el desarrollo cerebral de los niños.
El Proyecto Viva es un estudio prospectivo en curso que ha examinado la relación entre la ingesta diaria de nutrientes de donantes de metilo en 1,210 mujeres durante el embarazo y la cognición infantil a los tres y siete años después del parto. La ingesta materna de colina durante el primer y/o segundo trimestre del embarazo no se correlacionó con las medidas de rendimiento cognitivo en niños a los 3 años de edad (72). Otro reporte del estudio indicó que el cuartil superior versus inferior de las ingestas de colina materna durante el segundo trimestre del embarazo (ingestas medias, 392 mg/día versus 260 mg/día) se asoció significativamente con puntuaciones más altas de memoria visual en niños de 7 años de edad (73). Además, un pequeño ensayo aleatorizado, doble ciego, controlado con placebo en 99 mujeres embarazadas (de 21 a 41 años) evaluó el efecto de la suplementación con colina durante el embarazo y la lactancia sobre la función cognitiva de los bebés de 10 y 12 meses de edad (74). Los resultados indicaron que la suplementación materna con colina (750 mg/día de colina en la forma de fosfatidilcolina) desde las 18 semanas de gestación hasta los 3 meses posteriores al parto no proporcionó beneficios cognitivos en los niños con respecto a la memoria visoespacial a corto plazo, la memoria episódica a largo plazo, y el lenguaje y desarrollo global (74). En un estudio aleatorio, doble ciego, y de alimentación controlada en 24 mujeres embarazadas, la ingesta materna de colina de 930 mg/día (~2x la IA) durante el tercer trimestre del embarazo mejoró las medidas de velocidad de procesamiento de información (es decir, tiempos de reacción) y memoria visoespacial en bebés en comparación con ingestas de colina de 480 mg/día (75). La ingesta de colina en este estudio provino tanto de fuentes dietéticas como suplementarias: 380 mg/día de colina de fuentes dietéticas y 100 mg/día o 550 mg/día de cloruro de colina suplementario (75). Seguimiento de la descendencia (N=20) a los 7 años de edad indicó que los niños de madres que consumieron 930 mg/día de colina tenían mayores medidas de atención sostenida en comparación con los de madres que consumieron 480 mg/día de colina (76), lo que sugiere que la suplementación prenatal con colina durante la última etapa del embarazo puede conferir beneficios cognitivos para el niño.
Función cognitiva en adultos mayores
La función cognitiva, que incluye los dominios de memoria, velocidad, y función ejecutiva, disminuye gradualmente con el aumento de la edad. La tasa de deterioro cognitivo también está influenciada por factores de riesgo modificables como los hábitos alimenticios. La deficiencia de vitaminas B y las concentraciones elevadas de homocisteína en sangre se han asociado con deterioros cognitivos en los ancianos. Un meta-análisis reciente de 14 ensayos aleatorios controlados con placebo encontró que la suplementación con vitamina B desaceleró el deterioro cognitivo en adultos mayores cognitivamente sanos, medido por la puntuación en el Mini-Mental State Examination, en comparación con el placebo (77). Sin embargo, se sabe menos sobre la colina dietética o suplementaria específicamente.
Algunos estudios observacionales han examinado la ingesta de colina y la función cognitiva en adultos mayores, pero son de naturaleza de corte transversal. El análisis de datos de corte transversal de un subgrupo de 1,391 voluntarios (edades, 36-83 años) de la gran cohorte de Framingham Heart Study Offspring ha indicado que la ingesta dietética de colina se asoció positivamente con funciones cognitivas específicas, a saber, la memoria verbal y la memoria visual (78). En un estudio de corte transversal nacional de EE. UU. de 2,393 adultos mayores (≥60 años, las ingestas diarias totales de colina (combinada de la dieta y los suplementos) entre 187.06 y 399.5 mg/día se asociaron con un mejor rendimiento cognitivo en tres medidas separadas (evaluación del aprendizaje, velocidad de procesamiento, atención sostenida, y memoria de trabajo) en comparación con ingestas inferiores a 187.06 mg/día; sin embargo, las ingestas más altas de colina (>399.5 mg/día) no fueron diferentes de las ingestas más bajas (<187.06 mg/día) en estas medidas (79).
Otro estudio de corte transversal de 2,195 personas mayores (de 70 a 74 años) del Hordaland Health Study examinó las capacidades cognitivas y las concentraciones en la sangre de varios determinantes de la homocisteína circulante, incluidas la colina y la betaína (80). A diferencia de la betaína, las concentraciones en plasma altas versus bajas de colina libre (>8.36 µM frente a ≤8.36 µM) fueron significativamente asociadas con un mayor rendimiento en las pruebas cognitivas que evalúan la velocidad motora sensorial, la velocidad perceptiva, la función ejecutiva, y la cognición global. Sin embargo, en un estudio de intervención anterior en el que participaron 235 personas de edad avanzada (edad promedio, 81 años) con o sin deficiencia leve de vitamina B12, se encontró que las concentraciones basales de betaína — pero no de colina — estaban correlacionadas positivamente con las puntuaciones de las pruebas que evaluaban los los dominios cognitivos de construcción, velocidad sensorial del motor, y función ejecutiva (81).
Se necesita más investigación para determinar el efecto de la colina en el cerebro en desarrollo y si las ingestas de colina por encima de la IDR pueden ser útiles en la prevención de la pérdida de memoria o la demencia en humanos.
Tratamiento de Enfermedad
Enfermedades neurodegenerativas
Demencia
Las enfermedades neurodegenerativas, como la enfermedad de Alzheimer (EA) y la enfermedad de Parkinson (EP), se caracterizan por un deterioro cognitivo progresivo y demencia. Las disfunciones en la señalización de neurotransmisores, que afectan a las rutas colinérgicas y dopaminérgicas en particular, han estado involucradas en la aparición de deterioros cognitivos. Se han reportado déficits en la acetilcolina y un metabolismo anormal de los fosfolípidos en estudios posteriores a la muerte de los cerebros de pacientes con EA (12). Por estas razones, se han usado inhibidores de (acetil) colinesterasa (que cataliza la descomposición de la acetilcolina) y grandes dosis de lecitina (fosfatidilcolina) para tratar a pacientes con demencia debido a EA con la esperanza de aumentar la cantidad de acetilcolina disponible en el cerebro. Si bien los inhibidores de la colinesterasa han mostrado efectos positivos sobre las funciones cognitivas y las medidas del estado clínico global (82), una revisión sistemática de ensayos aleatorios controlados no encontró que la lecitina sea más beneficiosa que el placebo en el tratamiento de pacientes con deterioro cognitivo, demencia vascular, EA, o demencia mixta (83). Se dispone de datos limitados — en su mayoría de estudios de caso y control — para evaluar si la citicolina (CDP-colina) podría mejorar el rendimiento cognitivo en sujetos con EA o EP, y los estudios hasta la fecha generalmente han sido etiquetados como de mala calidad (84).
Glaucoma
Las neuropatías ópticas, incluido el glaucoma, se asocian con daño del nervio óptico y pérdida de la función visual. En el glaucoma, el deterioro progresivo del nervio óptico es causado por la pérdida de una población neuronal específica conocida como células ganglionares de la retina (CGR), de modo que la condición ha sido clasificada como una enfermedad neurodegenerativa (85). La colina y sus metabolitos desempeñan una serie de funciones para apoyar la función visual normal, incluida la función de la retina (86).
En un pequeño estudio, doble ciego, controlado con placebo, el efecto de la citicolina se evaluó en 24 sujetos afectados por glaucoma de ángulo abierto y tratados con β-bloqueadores. Los pacientes fueron aleatorizados para seguir un ciclo terapéutico durante un período total de ocho años: citicolina (1,000 mg/día, por inyección intramuscular) o placebo (solo β-bloqueadores) durante un período de dos meses seguido de un período de lavado de cuatro meses (87). Los exámenes electrofisiológicos se utilizaron para evaluar el alcance de las disfunciones visuales, incluidas las grabaciones simultáneas de ElectroRetinoGramas a Patrón (ERG-P) y Potenciales Evocados Visuales (PEV). Se encontró que la citicolina mejora la función de la retina y la conducción neural a lo largo de las rutas visuales post-retinianas, de modo que las respuestas de la corteza visual a los estímulos mejoraron significativamente en comparación con el placebo (87).
En un ensayo piloto similar, se evaluó la eficacia de la citicolina en 26 voluntarios (edad promedio, 65.4 años) afectados por otro tipo de neuropatía óptica conocida como neuropatía óptica isquémica anterior no arterítica (NOIANA). Se administró citicolina oral (1,600 mg/día) durante 60 días seguido de 60 días de lavado, y el ciclo terapéutico se repitió una sola vez. En comparación con placebo, se descubrió que la citicolina mejora la función de la retina y la conducción neural post-retiniana, evidenciada por las medidas de ERG-P y PEV (88). La citicolina oral (cuatro ciclos de 500 mg/día durante cuatro meses seguido de un período de lavado de dos meses) también fue asociada con la reducción significativa de la tasa de pérdida del campo visual y el nivel de presión intraocular en 41 pacientes con glaucoma progresivo (89). Se necesitan ensayos controlados aleatorios más grandes para establecer si la suplementación con citicolina podría incluirse en el tratamiento médico del glaucoma.
Algunos estudios también han explorado la citicolina tópica (es decir, gotas para los ojos) como un tratamiento potencial para el glaucoma. Un estudio en 56 pacientes con glaucoma de ángulo abierto encontró que la citicolina aplicada tópicamente durante cuatro meses mejoraba la función de la retina y la conducción neuronal relacionada con la visión cuando se combinaba con una monoterapia con betabloqueantes para reducir la presión intraocular (90). En un ensayo aleatorizado, doble ciego, y controlado con placebo en 78 pacientes con glaucoma progresivo de ángulo abierto (a pesar de presiones intraoculares de ≤18 mm Hg), el uso de gotas para los ojos de citicolina durante tres años mitigó la disminución del grosor de la capa de fibras nerviosas de la retina en comparación con el uso de gotas para los ojos con placebo (91). Este ensayo también encontró una tendencia (p=0.07) hacia una menor tasa de progresión de la enfermedad con citicolina tópica (91). Si bien los resultados de estos estudios piloto son prometedores, se necesitan ensayos controlados aleatorios a gran escala.
Enfermedades cerebrovasculares
Las enfermedades cerebrovasculares (incluidos los accidentes cerebrovasculares y la enfermedad cerebrovascular isquémica subaguda) son la principal causa de deterioro cognitivo en las personas mayores. Los resultados de estudios experimentales han sugerido que dosis farmacológicas de citicolina (CDP-colina) podrían mejorar el metabolismo de la glucosa y la biosíntesis de fosfolípidos y neurotransmisores, al tiempo que limitan la degradación de los fosfolípidos en las membranas neuronales en modelos de isquemia y enfermedades neurodegenerativas (revisado en 92). Muchos estudios de intervención a corto plazo en personas mayores con enfermedades vasculares han encontrado que las dosis terapéuticas de citicolina (administradas por vía oral, mediante inyección intramuscular, o mediante infusión intravenosa) produjeron mejoras en las funciones neuropsicológicas, incluidas las funciones cognitivas, emocionales, y conductuales (revisado en 5).
Un estudio observacional multicéntrico, de seis meses, inscribió a 197 sujetos con accidente cerebrovascular (edad promedio, 81.5 años) con un deterioro progresivo de su salud mental y confusión general y/o estupor a quienes inicialmente se les administró citicolina durante 5 o 10 días (2,000 mg/día, por infusión intravenosa) dentro de un período de cuatro meses, y luego durante 21 días (1,000 mg/día, por inyección intramuscular), repetido una vez después de un período de lavado de siete días (93). Se encontró que el tratamiento con citicolina se asocia con puntuaciones más altas en escalas de evaluación cognitiva y funcional en comparación con las mediciones iniciales. Sin embargo, sólo los ensayos controlados aleatorios podrían evaluar si la citicolina protege contra el daño vascular y el deterioro cognitivo en adultos mayores con síntomas geriátricos complejos.
El International Citicoline Trial on acUte Stroke (ICTUS) es un estudio multicéntrico y doble ciego que evaluó el efecto de suplementar a 2,298 pacientes con accidente cerebrovascular isquémico agudo con citicolina (2,000 mg/día) o un placebo durante seis semanas sobre varios resultados funcionales y neurológicos y en la tasa de mortalidad (94). Los resultados no mostraron diferencias entre los grupos de tratamiento después de un período de seguimiento de 90 días. Sólo los análisis de subgrupos encontraron beneficios significativos de la citicolina en pacientes mayores de 70 años, en aquellos con accidentes cerebrovasculares moderados en lugar de graves y en aquellos no tratados con activador del plasminógeno tisular recombinante (rtPA; tratamiento estándar de cuidado). Un meta-análisis anterior de pequeños ensayos aleatorios, controlados con placebo había informado un impacto positivo de la citicolina (1,000 mg/día, administrados durante 28 días a 12 meses) en la memoria y el comportamiento en sujetos con déficits cognitivos asociados con trastornos cerebrovasculares (95). El efecto de la citicolina también se evaluó en un ensayo multicéntrico, abierto, y controlado (ensayo IDEALE) en adultos mayores italianos (de 65 a 94 años) con evidencia de lesiones vasculares en neurorradiología y déficits cognitivos de leves a moderados, así como evaluado mediante el Mini-Examen del Estado Mental (MMSE; puntuaciones ≥21) (96). Trescientos cuarenta y nueve participantes recibieron citicolina oral (1,000 mg/día) o ningún tratamiento durante nueve meses. Las puntuaciones del MMSE en los individuos tratados con citicolina permanecieron sin cambios, mientras que se deterioraron significativamente en los pacientes no tratados, de modo que se encontró que las puntuaciones del MMSE entre los grupos eran significativamente diferentes después de tres y nueve meses de tratamiento. No se informó ningún efecto significativo en las medidas de autonomía funcional, estado de ánimo y trastornos del comportamiento. Otro ensayo abierto, aleatorizado, y controlado evaluó el efecto de la citicolina (1,000 mg/día durante 12 meses) en 347 sujetos (edad promedio, 67.2 años) que sufrieron un accidente cerebrovascular agudo. Los resultados demostraron que la citicolina limitó significativamente los deterioros cognitivos en los dominios de la atención y las funciones ejecutivas y la orientación temporal a los 6 y 12 meses después del accidente cerebrovascular en pacientes tratados en comparación con los no tratados (97). Sin embargo, otros ensayos aleatorios controlados no han encontrado ningún beneficio del tratamiento con citicolina en el tratamiento del accidente cerebrovascular isquémico agudo (98, 99), incluido un ensayo reciente que evaluó sus efectos cuando se administra inmediatamente después de la trombectomía endovascular (es decir, terapia de recanalización) y se continúa durante 42 días (100). Una revisión sistemática y un meta-análisis de 10 ensayos aleatorios controlados encontraron que el tratamiento con citicolina en el accidente cerebrovascular isquémico agudo (administrado por vía intravenosa y/u oral entre 8 horas y 14 días después del accidente cerebrovascular) se relacionó con una tasa ligeramente mayor de independencia (101). Sin embargo, un una revisión Cochrane reciente de ensayos controlados aleatorios encontró que la citicolina no tuvo ningún beneficio en los pacientes con accidente cerebrovascular isquémico agudo en comparación con el placebo con respecto a la mortalidad por todas las causas (8 ensayos); eventos cardiovasculares adversos, graves (3 ensayos); o grado de discapacidad o dependencia con las tareas diarias (4 ensayos) (102).
Por lo tanto, hay poca evidencia de que el tratamiento con citicolina sea eficaz en pacientes con enfermedad cerebrovascular. Muchos de los ensayos realizados no han tenido un diseño doble ciego, lo que reduce el riesgo de sesgo.
Lesión cerebral traumática
Durante décadas, estudios preclínicos y estudios clínicos pequeños han investigado el efecto de la citicolina en el tratamiento de pacientes que sufren una lesión cerebral traumática (LCT). Una revisión sistemática de datos clínicos realizada en 2011 sugirió que la citicolina podría acelerar la resorción del edema cerebral y mejorar la recuperación de la conciencia y los trastornos neurológicos en casos de LCT graves (clasificados según puntuaciones de ≤8 en la Escala de Coma de Glasgow [GCS]) (5). La citicolina también pareció limitar los déficits de memoria y la duración y gravedad de otros síntomas postraumáticos (p. ej., dolor de cabeza, mareos, trastorno de atención) en pacientes con LCT con lesiones leves a moderadas (puntuaciones GCS, 9-15) (revisado en 5).
Aunque la citicolina está incluida en el régimen terapéutico de LCT en 59 países, sólo se ha realizado un ensayo multicéntrico, aleatorizado, doble ciego, y controlado con placebo en los EE. UU. El Ensayo CiticOline Brain Injury (COBRIT) inscribió a 1,213 pacientes con LCT de leve a grave y evaluó el efecto de la citicolina enteral u oral (2,000 mg/día, durante 90 días) sobre los resultados funcionales y cognitivos (medidos por los componentes de la Batería de Cuidado de la Red de Ensayos Clínicos de LCT) (103). No se encontraron beneficios significativos de la suplementación con citicolina sobre el placebo a los 90 días (final del período de tratamiento) y a los 180 días. Es importante señalar que este ensayo tuvo una baja adherencia: solo el 44% de los pacientes en el ensayo tomaron al menos el 75% de la dosis asignada (103). Al combinar los resultados de COBRIT con los de otros 10 ensayos clínicos en un meta-análisis, el tratamiento con citicolina para el LCT en fase aguda se relacionó con un mayor grado de independencia, es decir, la capacidad de realizar actividades diarias sin necesidad de asistencia (104).
Fuentes
Síntesis de novo (biosíntesis)
Los humanos pueden sintetizar restos de colina en pequeñas cantidades convirtiendo fosfatidiletanolamina en fosfatidilcolina (ver Figura 2). Se requieren tres reacciones de metilación catalizadas por la fosfatidiletanolamina N-metiltransferasa (PEMT), cada una con S-adenosilmetionina (SAM) como donante del grupo metilo. La colina se genera de forma endógena cuando la metilación de fosfatidiletanolamina se combina con el catabolismo de fosfatidilcolina recién formada por fosfolipasas. Esto se conoce como síntesis de novo de colina. La sustitución de colina por serina en la síntesis de fosfatidilserina a partir de fosfatidilcolina por fosfatidilserina sintasa-1 también libera colina (4). Debido a que el metabolismo de fosfatidilcolina es una fuente de colina endógena, el nutriente no se clasificó inicialmente como esencial (1). Sin embargo, la síntesis de novo de colina en los humanos no son suficientes para satisfacer sus necesidades metabólicas, de modo que los humanos sanos alimentados con dietas deficientes en colina desarrollan hígado graso, daño hepático, y/o daño muscular (ver Deficiencia).
Fuentes alimenticias
En los EE. UU., las ingestas dietéticas promedio de colina están muy por debajo de la Ingesta Adecuada (IA) recomendada. Según una encuesta nacional de EE. UU., NHANES 2015-2018, la ingesta dietética promedio de colina fue de 284 mg/día para las mujeres y 390 mg/día para los hombres; sólo el 6% de las mujeres y el 11% de los hombres tenían ingestas dietéticas superiores a la IA (105). Además, los datos de NHANES indican que menos del 9% de las mujeres embarazadas cumplen con el IA de colina (106). Los estadounidenses de todas las edades tienen un consumo bajo de colina: un análisis de NHANES 2009-2012 encontró que menos del 11% de los residentes de EE. UU. de 2 años o más tenían una ingesta total de colina (de la dieta y los suplementos combinados) por encima de la IA (107). La estratificación de los datos por etapa de la vida indicó que los niños pequeños de 2 a 8 años años tenían más probabilidades de cumplir con la IA que los niños mayores, los adolescentes, o los adultos (107). Los vegetarianos, especialmente los veganos, que no consumen carne, leche ni huevos, pueden correr riesgo de una ingesta inadecuada de colina (26).
Los huevos, hígado, vegetales crucíferos, y cacahuetes son especialmente ricos en colina (32, 108); un análisis de los datos de NHANES concluyó que es extremadamente difícil alcanzar la IA únicamente a partir de fuentes alimentarias si no se consumen huevos (106). Los principales contribuyentes a la colina en la dieta estadounidense son la carne, las aves, el pescado, los productos lácteos, la pasta, el arroz, y los platos a base de huevo (109). Las espinacas, la remolacha, el trigo, y los mariscos también son buenas fuentes del metabolito de la colina, la betaína (110). La betaína no se puede volver a convertir en colina, pero puede ahorrar algunos requisitos de colina para la remetilación de la homocisteína (1).
La fosfatidilcolina, que contiene aproximadamente un 13% de colina por peso, es la principal forma de colina en los productos dietéticos (111). Los extractos de lecitina, que comprenden una mezcla de fosfatidilcolina y otros fosfolípidos, a menudo se añaden durante el procesamiento de alimentos. Se ha estimado que las lecitinas en los alimentos procesados aumentan el consumo diario de fosfatidilcolina en aproximadamente 1.5 mg/kg de peso corporal en los adultos (32).
El contenido total de colina de algunos alimentos que contienen colina se enumera en miligramos (mg) en la Tabla 2. Para obtener más información sobre el contenido de nutrientes de alimentos específicos, busque en la Base de Datos de los Alimentos de la USDA (FoodData Central) o la documentación del USDA sobre el contenido de colina de los alimentos comunes.
| Alimentos | Porción | Colina Total (mg) |
|---|---|---|
| Hígado de res, frito | 3 onzas* | 355 |
| Huevo | 1 grande | 151 |
| Vieira, cocida, al vapor | 3 onzas | 94 |
| Salmón, rosa, enlatado | 3 onzas | 75 |
| Carne de res, cortada en trozos, cocida | 3 onzas | 71 |
| Bacalao del Atlántico, cocido | 3 onzas | 71 |
| Camarones, enlatados | 3 onzas | 69 |
| Coles de Bruselas, cocidas, hervidas | 1 taza | 63 |
| Brócoli, cocido, hervido | 1 taza | 62 |
| Pollo, pechuga, cocido, asado | 3 onzas | 62 |
| Germen de trigo, tostado | ¼ taza | 51 |
| Leche, 1% | 1 taza | 43 |
| Frijoles lima, semillas inmaduras, cocidas | ½ taza | 34 |
| Mantequilla de maní, cremosa | 2 cucharadas | 20 |
| Maní | 1 onza | 15 |
| Almendras | 1 onza | 15 |
| *Una porción de 3 onzas de carne o pescado es aproximadamente del tamaño de un mazo de cartas. | ||
Suplementos
La CDP-colina (citicolina) y las sales de colina, como el cloruro de colina y el bitartrato de colina, están disponibles como suplementos. Los suplementos de fosfatidilcolina también aportan colina; sin embargo, la colina comprende sólo alrededor del 13% del peso de la fosfatidilcolina (111). Por lo tanto, un suplemento que contenga 4,230 mg (4.23 gramos) de fosfatidilcolina proporcionaría 550 mg de colina. Aunque el término "lecitina" es sinónimo de fosfatidilcolina cuando se usa en química, las lecitinas comerciales generalmente se preparan a partir de soja, girasol, y colza, y pueden contener entre 20%-90% de fosfatidilcolina. La lecitina de yema de huevo es una fuente menos probable de lecitina en los suplementos dietéticos. Además, la naturaleza de los ácidos grasos que contienen fosfatidilcolina depende de si la lecitina se produce a partir de fuentes vegetales, animales o microbianas. En particular, la lecitina de soja es más rica en ácidos grasos poliinsaturados que la lecitina de yema de huevo (112).
La mayoría de los suplementos multivitamínicos, incluidos los multivitamínicos prenatales, no contienen colina; los pocos que sí lo hacen suelen contener colina en niveles mucho más bajos que el IA (113).
Seguridad
Toxicidad
Las altas dosis (10,000 a 16,000 mg/día) de colina se han asociado con un olor corporal a pescado, vómitos, salivación, y aumento de la sudoración. El olor corporal a pescado resulta de la producción excesiva y la excreción de trimetilamina, un metabolito de la colina. En la condición hereditaria, la trimetilaminuria primaria (también conocida como "síndrome de olor a pescado"; ver el artículo sobre Riboflavina), una flavina defectuosa que contiene enzima monooxigenasa 3 (FMO3) da como resultado una oxidación deteriorada de la trimetilamina en el hígado. El manejo de la enfermedad incluye el uso de dietas restringidas en colina en las personas afectadas (114). La ingesta de grandes dosis de colina en forma de fosfatidilcolina (lecitina) generalmente no produce olor corporal a pescado, porque su metabolismo produce poca trimetilamina.
Se descubrió que una dosis de 7,500 mg/día de colina tenía un ligero efecto reductor de la presión arterial (hipotensor), que podía provocar mareos o desmayos. El trisalicilato de colina y magnesio en dosis de 3,000 mg/día ha provocado deterioro de la función hepática, picazón generalizada, y zumbidos en los oídos (tinnitus). Sin embargo, es probable que estos efectos fueran causados por el salicilato, más que por la colina en la preparación (32).
En 1998, la Junta de Alimentos y Nutrición (JNA) del Instituto de Medicina estableció el nivel máximo de ingesta tolerable (NM) para la colina en 3,500 mg/día para adultos (Tabla 3). Esta recomendación se basó principalmente en la prevención de la hipotensión (presión arterial baja) y, en segundo lugar, en la prevención del olor corporal a pescado debido al aumento de la excreción de trimetilamina. El NM se estableció para personas generalmente sanas, y el JNA señaló que las personas con enfermedad hepática o renal, enfermedad de Parkinson, depresión, o trimetilaminuria hereditaria podrían tener un mayor riesgo de efectos adversos al consumir colina a niveles cercanos al NM (32).
| Grupo Etario | NM (mg/día) |
|---|---|
| Infantes 0-12 meses | No es posible establecer* |
| Niños 1-8 años | 1,000 |
| Niños 9-13 años | 2,000 |
| Adolescentes 14-18 años | 3,000 |
| Adultos 19 años y más | 3,500 |
| *La fuente de la ingesta debe provenir sólo de alimentos y fórmula. | |
¿Las ingestas altas de colina y/o los suplementos de fosfatidilcolina aumentan el riesgo de enfermedad cardiovascular?
Se ha encontrado que la suplementación oral con fosfatidilcolina (250 mg de colina total de los alimentos más 250 mg de fosfatidilcolina suplementaria) produce concentraciones detectables de trimetilamina y N-óxido de trimetilamina (TMAO) en la sangre (23). La microbiota intestinal está directamente implicada en la generación de trimetilamina a partir de colina dietética y su metabolito fosfatidilcolina, así como de betaína y carnitina dietética. Posteriormente, la trimetilamina se convierte en TMAO mediante monooxigenasas que contienen flavina en el hígado. El estudio prospectivo que siguió a 4,007 personas — con o sin enfermedad cardiovascular — durante un período de tres años encontró que las concentraciones basales de TMAO circulante se correlacionaban positivamente con la incidencia de muerte, infarto al miocardio no fatal y accidente cerebrovascular, descritos como eventos adversos cardiovasculares mayores (MACE) (23). En la misma cohorte, se encontró que el riesgo de MACE era aproximadamente un 30% mayor en individuos en el cuartil más alto versus el más bajo de concentraciones en plasma de colina o betaína (115). Sin embargo, dependiendo de la composición de la microbiota intestinal, el riesgo de tener un evento adverso cardiovascular puede ser menor en individuos con TMAO circulante bajo versus alto, incluso aunque las concentraciones de colina y/o betaína en la sangre estén elevadas (115). Desde entonces se han realizado hallazgos similares en otras cohortes prospectivas. Un meta-análisis de 19 estudios prospectivos encontró que las concentraciones más altas de TMAO en sangre se asociaron con un 62% más de riesgo de MACE o muerte en comparación con concentraciones más bajas (116). Las concentraciones elevadas de TMAO circulante no solo se han relacionado con un mayor riesgo de enfermedad cardiovascular sino también a un mayor riesgo de diabetes mellitus tipo 2 y enfermedad renal (117-119). Sin embargo, las concentraciones elevadas de TMAO en la sangre pueden ser un biomarcador de la enfermedad más que un factor causante (120, 121).
Se necesitan más investigaciones para comprender cómo la composición de la microbiota intestinal influye en el destino metabólico de la colina ingerida. En la actualidad, hay poca evidencia de que la colina dietética aumente el riesgo de eventos cardiovasculares. Los estudios prospectivos de cohortes sobre la asociación han sido inconsistentes: algunos estudios han relacionado una mayor ingesta de colina con mayores riesgos de fibrilación auricular (122) y mortalidad relacionada con enfermedades cardiovasculares (43, 44), mientras que varios otros estudios prospectivos no han encontrado asociación entre la ingesta de colina y criterios de evaluación de enfermedades cardiovasculares (41, 45-48). Curiosamente, en el ensayo PREvención con DIeta MEDiterránea-Plus (Predimed-Plus) en personas con sobrepeso u obesidad con síndrome metabólico, un aumento en la ingesta de colina dietética durante un período de un año se asoció con mejores medidas de salud cardiovascular (es decir, colesterol total sérico menor, el colesterol LDL sérico, y los niveles de presión arterial diastólica y sistólica) (123).
Interacción con drogas/fármacos
El metotrexato, un medicamento utilizado en el tratamiento del cáncer, la psoriasis, y la artritis reumatoide, inhibe la enzima dihidrofolato reductasa y, por lo tanto, limita la disponibilidad de grupos metilo donados a partir de derivados de folato. Las ratas que recibieron metotrexato han mostrado evidencia de disminución del estado nutricional de la colina y mayores reacciones adversas a los medicamentos debido a la disfunción hepática (12, 124). Por lo tanto, las personas que toman metotrexato pueden tener un mayor requerimiento de colina. Los tratamientos con una familia de fármacos hipolipemiantes conocidos como fibratos (p. ej., fenofibrato, bezofibrato) se han asociado con una mayor excreción de betaína en la orina y un aumento de la concentración de homocisteína en la sangre de pacientes con diabetes mellitus o síndrome metabólico (125, 126). Si los beneficios de la terapia de fibrato se mitigan de hecho por la deficiencia de betaína inducida por fibrato, el uso y la seguridad de suplementar a los pacientes con betaína deberían considerarse (127).
Recomendación del LPI
Poco se sabe sobre la cantidad de colina en la dieta requerida para promover una salud óptima o prevenir enfermedades crónicas en humanos. El Instituto Linus Pauling (LPI) apoya la recomendación de la Junta de Alimentos y Nutrición de 425 mg/día para mujeres adultas y 550 mg/día para hombres adultos. Una dieta saludable, y variada debería proporcionar suficiente colina para la mayoría de las personas, pero los vegetarianos estrictos que no consumen leche ni huevos pueden correr el riesgo de una ingesta inadecuada de colina.
Adultos mayores (>50 años)
Poco se sabe sobre la cantidad de colina dietética con mayor probabilidad de promover una salud óptima o prevenir enfermedades crónicas en adultos mayores. En la actualidad, no hay evidencia que respalde una ingesta recomendada diferente de colina de la de los adultos más jóvenes (425 mg/día para las mujeres y 550 mg/día para los hombres).
Autores y Críticos
Originalmente escrito en Noviembre 2003 por:
Jane Higdon, Ph.D.
Instituto Linus Pauling
Universidad Estatal de Oregon
Actualizado en Enero 2015 por:
Barbara Delage, Ph.D.
Instituto Linus Pauling
Universidad Estatal de Oregon
Actualizado en Abril 2023 by:
Victoria J. Drake, Ph.D.
Instituto Linus Pauling
Universidad Estatal de Oregon
Revisado en Julio 2023 por:
Isis Trujillo-Gonzalez, Ph.D.
Profesor Asistente
Instituto de Investigación en Nutrición
La Universidad de Carolina del Norte, Chapel Hill
Derechos de autoría 2000-2026 Instituto Linus Pauling
La traducción de este artículo fue financiada por una subvención de Balchem Corporation.
Figure Alternative Text
Figure 1. Chemical structures of choline and derivatives
The figure shows the chemical structures of choline, acetylcholine, betaine (trimethylglycine), glycerophosphorylcholine, sphingomyelin, and phosphatidylcholine.
Figure 2a. Synthesis of phosphatidylcholine
The figure illustrates the cytidine diphosphate (CDP)-choline pathway for phosphatidylcholine synthesis, showing chemical structures of intermediates and involved enzymes. At the top, choline is phosphorylated by choline kinase using ATP and releasing ADP. Phosphocholine is then converted to CDP-choline by CTP:choline phosphate cytidylyltransferase, using CTP and releasing pyrophosphate (PPi). Finally, CDP-choline reacts with diacylglycerol (DAG) or alkyl-acylglycerol (AAG) via cytidine diphosphocholine:1,2-diacylglycerol choline phosphotransferase to form phosphatidylcholine, releasing cytidine monophosphate (CMP).
Figure 2b. Synthesis of phosphatidylcholine
The figure shows the methylation pathway for phosphatidylcholine synthesis, including chemical structures. Phosphatidylethanolamine is converted to phosphatidylcholine by phosphatidylethanolamine N-methyltransferase. This reaction uses three molecules of S-adenosylmethionine (SAM) as methyl donors and releases three molecules of S-adenosylhomocysteine (SAH).
Figure 3. Synthesis of sphingomyelin
The figure illustrates the pathway of sphingomyelin synthesis. Sphingomyelin synthase catalyzes the transfer of a phosphocholine headgroup from phosphatidylcholine to ceramide, generating sphingomyelin and 1,2-diacylglycerol.
Figure 4. Homocysteine metabolism
The figure shows pathways of homocysteine metabolism in two panels:
Panel A: Homocysteine is methylated to form the essential amino acid methionine in two pathways. The first pathway involves remethylation catalyzed by the vitamin B12-dependent methionine synthase, which captures a methyl group from the folate-dependent one-carbon pool (5-methyltetrahydrofolate). The second pathway uses betaine as a methyl donor, catalyzed by betaine homocysteine methyltransferase (BHMT).
The catabolic transsulfuration pathway converts homocysteine to cysteine via two vitamin B6-dependent enzymes: cystathionine β synthase catalyzes the condensation of homocysteine with serine to form cystathionine, and cystathionine γ lyase converts cystathionine to cysteine, α-ketobutyrate, and ammonia.
Panel B: Methionine is the precursor of the universal methyl donor, S-adenosylmethionine (SAM). Three SAM molecules are required for the methylation of phosphatidylethanolamine to phosphatidylcholine by phosphatidylethanolamine N-methyltransferase (PEMT). Choline can be generated from phosphatidylcholine via phospholipases or converted to phosphatidylcholine via the cytidine diphospho-choline pathway (see Figure 2a).
Referencias
- Zeisel SH, Corbin KD. Choline. Present Knowledge in Nutrition. 10th ed: John Wiley & Sons, Inc.; 2012:405-418.
- Ueland PM. Choline and betaine in health and disease. J Inherit Metab Dis. 2011;34(1):3-15. (PubMed)
- Gibellini F, Smith TK. The Kennedy pathway--De novo synthesis of phosphatidylethanolamine and phosphatidylcholine. IUBMB Life. 2010;62(6):414-428. (PubMed)
- Li Z, Vance DE. Phosphatidylcholine and choline homeostasis. J Lipid Res. 2008;49(6):1187-1194. (PubMed)
- Secades JJ. Citicoline: pharmacological and clinical review, 2010 update. Rev Neurol. 2011;52 Suppl 2:S1-S62. (PubMed)
- Beckmann J, Lips KS. The non-neuronal cholinergic system in health and disease. Pharmacology. 2013;92(5-6):286-302. (PubMed)
- Noga AA, Vance DE. A gender-specific role for phosphatidylethanolamine N-methyltransferase-derived phosphatidylcholine in the regulation of plasma high density and very low density lipoproteins in mice. J Biol Chem. 2003;278(24):21851-21859. (PubMed)
- Noga AA, Zhao Y, Vance DE. An unexpected requirement for phosphatidylethanolamine N-methyltransferase in the secretion of very low density lipoproteins. J Biol Chem. 2002;277(44):42358-42365. (PubMed)
- da Costa KA, Kozyreva OG, Song J, Galanko JA, Fischer LM, Zeisel SH. Common genetic polymorphisms affect the human requirement for the nutrient choline. Faseb J. 2006;20(9):1336-1344. (PubMed)
- Pellanda H. Betaine homocysteine methyltransferase (BHMT)-dependent remethylation pathway in human healthy and tumoral liver. Clin Chem Lab Med. 2013;51(3):617-621. (PubMed)
- Gerhard GT, Duell PB. Homocysteine and atherosclerosis. Curr Opin Lipidol. 1999;10(5):417-428. (PubMed)
- Zeisel SH. Choline. In: Ross A, Caballero B, Cousins R, Tucker K, Ziegler T, eds. Modern Nutrition in Health and Disease. 11th ed: Lippincott Williams & Wilkins; 2014:416-426.
- Michelotti GA, Machado MV, Diehl AM. NAFLD, NASH and liver cancer. Nat Rev Gastroenterol Hepatol. 2013;10(11):656-665. (PubMed)
- Zeisel SH, Blusztajn JK. Choline and human nutrition. Annu Rev Nutr. 1994;14:269-296. (PubMed)
- da Costa KA, Niculescu MD, Craciunescu CN, Fischer LM, Zeisel SH. Choline deficiency increases lymphocyte apoptosis and DNA damage in humans. Am J Clin Nutr. 2006;84(1):88-94. (PubMed)
- Rolo AP, Teodoro JS, Palmeira CM. Role of oxidative stress in the pathogenesis of nonalcoholic steatohepatitis. Free Radic Biol Med. 2012;52(1):59-69. (PubMed)
- Fischer LM, daCosta KA, Kwock L, et al. Sex and menopausal status influence human dietary requirements for the nutrient choline. Am J Clin Nutr. 2007;85(5):1275-1285. (PubMed)
- Fischer LM, da Costa KA, Kwock L, Galanko J, Zeisel SH. Dietary choline requirements of women: effects of estrogen and genetic variation. Am J Clin Nutr. 2010;92(5):1113-1119. (PubMed)
- Resseguie M, Song J, Niculescu MD, da Costa KA, Randall TA, Zeisel SH. Phosphatidylethanolamine N-methyltransferase (PEMT) gene expression is induced by estrogen in human and mouse primary hepatocytes. Faseb J. 2007;21(10):2622-2632. (PubMed)
- da Costa KA, Corbin KD, Niculescu MD, Galanko JA, Zeisel SH. Identification of new genetic polymorphisms that alter the dietary requirement for choline and vary in their distribution across ethnic and racial groups. FASEB J. 2014;28(7):2970-2978. (PubMed)
- Kohlmeier M, da Costa KA, Fischer LM, Zeisel SH. Genetic variation of folate-mediated one-carbon transfer pathway predicts susceptibility to choline deficiency in humans. Proc Natl Acad Sci U S A. 2005;102(44):16025-16030. (PubMed)
- Spencer MD, Hamp TJ, Reid RW, Fischer LM, Zeisel SH, Fodor AA. Association between composition of the human gastrointestinal microbiome and development of fatty liver with choline deficiency. Gastroenterology. 2011;140(3):976-986. (PubMed)
- Tang WH, Wang Z, Levison BS, et al. Intestinal microbial metabolism of phosphatidylcholine and cardiovascular risk. N Engl J Med. 2013;368(17):1575-1584. (PubMed)
- Wang Z, Klipfell E, Bennett BJ, et al. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature. 2011;472(7341):57-63. (PubMed)
- Korsmo HW, Jiang X, Caudill MA. Choline: exploring the growing science on its benefits for moms and babies. Nutrients. 2019;11(8). (PubMed)
- Wallace TC, Blusztajn JK, Caudill MA, et al. Choline: the underconsumed and underappreciated essential nutrient. Nutr Today. 2018;53(6):240-253. (PubMed)
- Bernhard W, Shunova A, Machann J, et al. Resolution of severe hepatosteatosis in a cystic fibrosis patient with multifactorial choline deficiency: A case report. Nutrition. 2021;89:111348. (PubMed)
- Grothe J, Riethmuller J, Tschurtz SM, et al. Plasma phosphatidylcholine alterations in cystic fibrosis patients: impaired metabolism and correlation with lung function and inflammation. Cell Physiol Biochem. 2015;35(4):1437-1453. (PubMed)
- Bernhard W, Poets CF, Franz AR. Choline and choline-related nutrients in regular and preterm infant growth. Eur J Nutr. 2019;58(3):931-945. (PubMed)
- Jacob RA, Jenden DJ, Allman-Farinelli MA, Swendseid ME. Folate nutriture alters choline status of women and men fed low choline diets. J Nutr. 1999;129(3):712-717. (PubMed)
- Kim YI, Miller JW, da Costa KA, et al. Severe folate deficiency causes secondary depletion of choline and phosphocholine in rat liver. J Nutr. 1994;124(11):2197-2203. (PubMed)
- Food and Nutrition Board, Institute of Medicine. Choline. Dietary referencia Intakes: Thiamin, Riboflavin, Niacin, Vitamin B-6, Vitamin B-12, Pantothenic Acid, Biotin, and Choline. Washington D.C.: National Academy Press; 1998:390-422. (National Academy Press)
- Ganz AB, Klatt KC, Caudill MA. Common genetic variants alter metabolism and influence dietary choline requirements. Nutrients. 2017;9(8):252. (PubMed)
- Ganz AB, Cohen VV, Swersky CC, et al. Genetic variation in choline-metabolizing enzymes alters choline metabolism in young women consuming choline intakes meeting current recommendations. Int J Mol Sci. 2017;18(2):837. (PubMed)
- Zhou J, Austin RC. Contributions of hyperhomocysteinemia to atherosclerosis: Causal relationship and potential mechanisms. Biofactors. 2009;35(2):120-129. (PubMed)
- Leach NV, Dronca E, Vesa SC, et al. Serum homocysteine levels, oxidative stress and cardiovascular risk in non-alcoholic steatohepatitis. Eur J Intern Med. 2014;25(8):762-767. (PubMed)
- Olthof MR, Brink EJ, Katan MB, Verhoef P. Choline supplemented as phosphatidylcholine decreases fasting and postmethionine-loading plasma homocysteine concentrations in healthy men. Am J Clin Nutr. 2005;82(1):111-117. (PubMed)
- Olthof MR, van Vliet T, Boelsma E, Verhoef P. Low dose betaine supplementation leads to immediate and long term lowering of plasma homocysteine in healthy men and women. J Nutr. 2003;133(12):4135-4138. (PubMed)
- Schwab U, Torronen A, Toppinen L, et al. Betaine supplementation decreases plasma homocysteine concentrations but does not affect body weight, body composition, or resting energy expenditure in human subjects. Am J Clin Nutr. 2002;76(5):961-967. (PubMed)
- Steenge GR, Verhoef P, Katan MB. Betaine supplementation lowers plasma homocysteine in healthy men and women. J Nutr. 2003;133(5):1291-1295. (PubMed)
- Dalmeijer GW, Olthof MR, Verhoef P, Bots ML, van der Schouw YT. Prospective study on dietary intakes of folate, betaine, and choline and cardiovascular disease risk in women. Eur J Clin Nutr. 2008;62(3):386-394. (PubMed)
- Millard HR, Musani SK, Dibaba DT, et al. Dietary choline and betaine; associations with subclinical markers of cardiovascular disease risk and incidence of CVD, coronary heart disease and stroke: the Jackson Heart Study. Eur J Nutr. 2018;57(1):51-60. (PubMed)
- Zheng Y, Li Y, Rimm EB, et al. Dietary phosphatidylcholine and risk of all-cause and cardiovascular-specific mortality among US women and men. Am J Clin Nutr. 2016;104(1):173-180. (PubMed)
- Yang JJ, Lipworth LP, Shu XO, et al. Associations of choline-related nutrients with cardiometabolic and all-cause mortality: results from 3 prospective cohort studies of blacks, whites, and Chinese. Am J Clin Nutr. 2020;111(3):644-656. (PubMed)
- Nagata C, Wada K, Tamura T, et al. Choline and betaine intakes are not associated with cardiovascular disease mortality risk in Japanese men and women. J Nutr. 2015;145(8):1787-1792. (PubMed)
- Bidulescu A, Chambless LE, Siega-Riz AM, Zeisel SH, Heiss G. Usual choline and betaine dietary intake and incident coronary heart disease: the Atherosclerosis Risk in Communities (ARIC) study. BMC Cardiovasc Disord. 2007;7:20. (PubMed)
- Bertoia ML, Pai JK, Cooke JP, et al. Plasma homocysteine, dietary B vitamins, betaine, and choline and risk of peripheral artery disease. Atherosclerosis. 2014;235(1):94-101. (PubMed)
- Golzarand M, Mirmiran P, Azizi F. Association between dietary choline and betaine intake and 10.6-year cardiovascular disease in adults. Nutr J. 2022;21(1):1. (PubMed)
- Lundberg P, Dudman NP, Kuchel PW, Wilcken DE. 1H NMR determination of urinary betaine in patients with premature vascular disease and mild homocysteinemia. Clin Chem. 1995;41(2):275-283. (PubMed)
- Lever M, George PM, Elmslie JL, et al. Betaine and secondary events in an acute coronary syndrome cohort. PLoS One. 2012;7(5):e37883. (PubMed)
- Konstantinova SV, Tell GS, Vollset SE, Nygard O, Bleie O, Ueland PM. Divergent associations of plasma choline and betaine with components of metabolic syndrome in middle age and elderly men and women. J Nutr. 2008;138(5):914-920. (PubMed)
- Meyer KA, Benton TZ, Bennett BJ, et al. Microbiota-dependent metabolite trimethylamine N-oxide and coronary artery calcium in the coronary artery risk development in young adults study (CARDIA). J Am Heart Assoc. 2016;5(10). (PubMed)
- Yu D, Shu XO, Xiang YB, et al. Higher dietary choline intake is associated with lower risk of nonalcoholic fatty liver in normal-weight Chinese women. J Nutr. 2014;144(12):2034-2040. (PubMed)
- Guerrerio AL, Colvin RM, Schwartz AK, et al. Choline intake in a large cohort of patients with nonalcoholic fatty liver disease. Am J Clin Nutr. 2012;95(4):892-900. (PubMed)
- Mazidi M, Katsiki N, Mikhailidis DP, Banach M. Adiposity may moderate the link between choline intake and non-alcoholic fatty liver disease. J Am Coll Nutr. 2019;38(7):633-639. (PubMed)
- Talaulikar VS, Arulkumaran S. Folic acid in obstetric practice: a review. Obstet Gynecol Surv. 2011;66(4):240-247. (PubMed)
- Czeizel AE, Dudas I, Vereczkey A, Banhidy F. Folate deficiency and folic acid supplementation: the prevention of neural-tube defects and congenital heart defects. Nutrients. 2013;5(11):4760-4775. (PubMed)
- Eskes TK. Open or closed? A world of difference: a history of homocysteine research. Nutr Rev. 1998;56(8):236-244. (PubMed)
- Shaw GM, Carmichael SL, Yang W, Selvin S, Schaffer DM. Periconceptional dietary intake of choline and betaine and neural tube defects in offspring. Am J Epidemiol. 2004;160(2):102-109. (PubMed)
- Carmichael SL, Yang W, Shaw GM. Periconceptional nutrient intakes and risks of neural tube defects in California. Birth Defects Res A Clin Mol Teratol. 2010;88(8):670-678. (PubMed)
- Chandler AL, Hobbs CA, Mosley BS, et al. Neural tube defects and maternal intake of micronutrients related to one-carbon metabolism or antioxidant activity. Birth Defects Res A Clin Mol Teratol. 2012;94(11):864-874. (PubMed)
- Petersen JM, Parker SE, Crider KS, Tinker SC, Mitchell AA, Werler MM. One-carbon cofactor intake and risk of neural tube defects among women who meet folic acid recommendations: a multicenter case-control study. Am J Epidemiol. 2019;188(6):1136-1143. (PubMed)
- Lavery AM, Brender JD, Zhao H, et al. Dietary intake of choline and neural tube defects in Mexican Americans. Birth Defects Res A Clin Mol Teratol. 2014;100(6):463-471. (PubMed)
- Shaw GM, Finnell RH, Blom HJ, et al. Choline and risk of neural tube defects in a folate-fortified population. Epidemiology. 2009;20(5):714-719. (PubMed)
- Mills JL, Fan R, Brody LC, et al. Maternal choline concentrations during pregnancy and choline-related genetic variants as risk factors for neural tube defects. Am J Clin Nutr. 2014;100(4):1069-1074. (PubMed)
- Obeid R, Derbyshire E, Schon C. Association between maternal choline, fetal brain development, and child neurocognition: systematic review and meta-analysis of human studies. Adv Nutr. 2022;13(6):2445-2457. (PubMed)
- Teather LA, Wurtman RJ. Dietary cytidine (5')-diphosphocholine supplementation protects against development of memory deficits in aging rats. Prog Neuropsychopharmacol Biol Psychiatry. 2003;27(4):711-717. (PubMed)
- Zeisel SH. Choline: critical role during fetal development and dietary requirements in adults. Annu Rev Nutr. 2006;26:229-250. (PubMed)
- McCann JC, Hudes M, Ames BN. An overview of evidence for a causal relationship between dietary availability of choline during development and cognitive function in offspring. Neurosci Biobehav Rev. 2006;30(5):696-712. (PubMed)
- Dauncey MJ. Nutrition, the brain and cognitive decline: insights from epigenetics. Eur J Clin Nutr. 2014;68(11):1179-1185. (PubMed)
- Strain JJ, McSorley EM, van Wijngaarden E, et al. Choline status and neurodevelopmental outcomes at 5 years of age in the Seychelles Child Development Nutrition Study. Br J Nutr. 2013;110(2):330-336. (PubMed)
- Villamor E, Rifas-Shiman SL, Gillman MW, Oken E. Maternal intake of methyl-donor nutrients and child cognition at 3 years of age. Paediatr Perinat Epidemiol. 2012;26(4):328-335. (PubMed)
- Boeke CE, Gillman MW, Hughes MD, Rifas-Shiman SL, Villamor E, Oken E. Choline intake during pregnancy and child cognition at age 7 years. Am J Epidemiol. 2013;177(12):1338-1347. (PubMed)
- Cheatham CL, Goldman BD, Fischer LM, da Costa KA, Reznick JS, Zeisel SH. Phosphatidylcholine supplementation in pregnant women consuming moderate-choline diets does not enhance infant cognitive function: a randomized, double-blind, placebo-controlled trial. Am J Clin Nutr. 2012;96(6):1465-1472. (PubMed)
- Caudill MA, Strupp BJ, Muscalu L, Nevins JEH, Canfield RL. Maternal choline supplementation during the third trimester of pregnancy improves infant information processing speed: a randomized, double-blind, controlled feeding study. FASEB J. 2018;32(4):2172-2180. (PubMed)
- Bahnfleth CL, Strupp BJ, Caudill MA, Canfield RL. Prenatal choline supplementation improves child sustained attention: A 7-year follow-up of a randomized controlled feeding trial. FASEB J. 2022;36(1):e22054. (PubMed)
- Wang Z, Zhu W, Xing Y, Jia J, Tang Y. B vitamins and prevention of cognitive decline and incident dementia: a systematic review and meta-analysis. Nutr Rev. 2022;80(4):931-949. (PubMed)
- Poly C, Massaro JM, Seshadri S, et al. The relation of dietary choline to cognitive performance and white-matter hyperintensity in the Framingham Offspring Cohort. Am J Clin Nutr. 2011;94(6):1584-1591. (PubMed)
- Liu L, Qiao S, Zhuang L, et al. Choline intake correlates with cognitive performance among elder adults in the United States. Behav Neurol. 2021;2021:2962245. (PubMed)
- Nurk E, Refsum H, Bjelland I, et al. Plasma free choline, betaine and cognitive performance: the Hordaland Health Study. Br J Nutr. 2013;109(3):511-519. (PubMed)
- Eussen SJ, Ueland PM, Clarke R, et al. The association of betaine, homocysteine and related metabolites with cognitive function in Dutch elderly people. Br J Nutr. 2007;98(5):960-968. (PubMed)
- Birks J. Cholinesterase inhibitors for Alzheimer's disease. Cochrane Database Syst Rev. 2006(1):CD005593. (PubMed)
- Higgins JP, Flicker L. Lecithin for dementia and cognitive impairment. Cochrane Database Syst Rev. 2003(3):CD001015. (PubMed)
- Bonvicini M, Travaglini S, Lelli D, Antonelli Incalzi R, Pedone C. Is citicoline effective in preventing and slowing down dementia?-a systematic review and a meta-analysis. Nutrients. 2023;15(2):386. (PubMed)
- Gupta N, Yucel YH. Glaucoma as a neurodegenerative disease. Curr Opin Ophthalmol. 2007;18(2):110-114. (PubMed)
- Hwang JS, Shin YJ. Role of choline in ocular diseases. Int J Mol Sci. 2021;22(9):4733. (PubMed)
- Parisi V. Electrophysiological assessment of glaucomatous visual dysfunction during treatment with cytidine-5'-diphosphocholine (citicoline): a study of 8 years of follow-up. Doc Ophthalmol. 2005;110(1):91-102. (PubMed)
- Parisi V, Coppola G, Ziccardi L, Gallinaro G, Falsini B. Cytidine-5'-diphosphocholine (Citicoline): a pilot study in patients with non-arteritic ischaemic optic neuropathy. Eur J Neurol. 2008;15(5):465-474. (PubMed)
- Ottobelli L, Manni GL, Centofanti M, Iester M, Allevena F, Rossetti L. Citicoline oral solution in glaucoma: is there a role in slowing disease progression? Ophthalmologica. 2013;229(4):219-226. (PubMed)
- Parisi V, Centofanti M, Ziccardi L, et al. Treatment with citicoline eye drops enhances retinal function and neural conduction along the visual pathways in open angle glaucoma. Graefes Arch Clin Exp Ophthalmol. 2015;253(8):1327-1340. (PubMed)
- Rossetti L, Iester M, Tranchina L, et al. Can treatment with citicoline eyedrops reduce progression in glaucoma? The results of a randomized placebo-controlled clinical trial. J Glaucoma. 2020;29(7):513-520. (PubMed)
- Grieb P. Neuroprotective properties of citicoline: facts, doubts and unresolved issues. CNS Drugs. 2014;28(3):185-193. (PubMed)
- Putignano S, Gareri P, Castagna A, et al. Retrospective and observational study to assess the efficacy of citicoline in elderly patients suffering from stupor related to complex geriatric syndrome. Clin Interv Aging. 2012;7:113-118. (PubMed)
- Davalos A, Alvarez-Sabin J, Castillo J, et al. Citicoline in the treatment of acute ischaemic stroke: an international, randomised, multicentre, placebo-controlled study (ICTUS trial). Lancet. 2012;380(9839):349-357. (PubMed)
- Fioravanti M, Yanagi M. Cytidinediphosphocholine (CDP-choline) for cognitive and behavioural disturbances associated with chronic cerebral disorders in the elderly. Cochrane Database Syst Rev. 2005(2):CD000269. (PubMed)
- Cotroneo AM, Castagna A, Putignano S, et al. Effectiveness and safety of citicoline in mild vascular cognitive impairment: the IDEALE study. Clin Interv Aging. 2013;8:131-137. (PubMed)
- Alvarez-Sabin J, Ortega G, Jacas C, et al. Long-term treatment with citicoline may improve poststroke vascular cognitive impairment. Cerebrovasc Dis. 2013;35(2):146-154. (PubMed)
- Clark WM, Williams BJ, Selzer KA, Zweifler RM, Sabounjian LA, Gammans RE. A randomized efficacy trial of citicoline in patients with acute ischemic stroke. Stroke. 1999;30(12):2592-2597. (PubMed)
- Clark WM, Wechsler LR, Sabounjian LA, Schwiderski UE, Citicoline Stroke Study Group. A phase III randomized efficacy trial of 2000 mg citicoline in acute ischemic stroke patients. Neurology. 2001;57(9):1595-1602. (PubMed)
- Agarwal A, Vishnu VY, Sharma J, et al. Citicoline in acute ischemic stroke: A randomized controlled trial. PLoS One. 2022;17(5):e0269224. (PubMed)
- Secades JJ, Alvarez-Sabin J, Castillo J, et al. Citicoline for acute ischemic atroke: a systematic review and formal meta-analysis of randomized, double-blind, and placebo-controlled trials. J Stroke Cerebrovasc Dis. 2016;25(8):1984-1996. (PubMed)
- Marti-Carvajal AJ, Valli C, Marti-Amarista CE, Sola I, Marti-Fabregas J, Bonfill Cosp X. Citicoline for treating people with acute ischemic stroke. Cochrane Database Syst Rev. 2020;8(8):CD013066. (PubMed)
- Zafonte RD, Bagiella E, Ansel BM, et al. Effect of citicoline on functional and cognitive status among patients with traumatic brain injury: Citicoline Brain Injury Treatment Trial (COBRIT). JAMA. 2012;308(19):1993-2000. (PubMed)
- Secades JJ, Trimmel H, Salazar B, Gonzalez JA. Citicoline for the management of patients with traumatic brain injury in the acute phase: a systematic review and meta-analysis. Life (Basel). 2023;13(2):369. (PubMed)
- USDA, Agricultural Research Service, 2021. Usual Nutrient Intake from Food and Beverages, by Gender and Age, What We Eat in America, NHANES 2015-2018. Available http://www.ars.usda.gov/nea/bhnrc/fsrg.
- Wallace TC, Fulgoni VL 3rd. Usual choline intakes are associated with egg and protein food consumption in the United States. Nutrients. 2017;9(8):839. (PubMed)
- Wallace TC, Fulgoni VL, 3rd. Assessment of total choline intakes in the United States. J Am Coll Nutr. 2016;35(2):108-112. (PubMed)
- Zeisel SH, Klatt KC, Caudill MA. Choline. Adv Nutr. 2018;9(1):58-60. (PubMed)
- Chester DN, Goldman JD, Ahuja JK, Moshfegh AJ. Dietary Intakes of Choline: What We Eat in America, NHANES 2007-2008. Food Surveys Research Group Dietary Data Brief No. 9. October 2011. Available at: http://ars.usda.gov/Services/docs.htm?docid=19476.
- Craig SA. Betaine in human nutrition. Am J Clin Nutr. 2004;80(3):539-549. (PubMed)
- Koc H, Mar MH, Ranasinghe A, Swenberg JA, Zeisel SH. Quantitation of choline and its metabolites in tissues and foods by liquid chromatography/electrospray ionization-isotope dilution mass spectrometry. Anal Chem. 2002;74(18):4734-4740. (PubMed)
- Hendler SS, Rorvik DR, eds. PDR for Nutritional Supplements. 2nd ed: Thomson Reuters; 2008.
- US Department of Health and Human Services, National Institutes of Health, Office of Dietary Supplements. Dietary Supplement Label Database (DSLD). [Internet]. [cited 4/10/2023]. Available from: https://dsld.od.nih.gov/.
- Busby MG, Fischer L, da Costa KA, Thompson D, Mar MH, Zeisel SH. Choline- and betaine-defined diets for use in clinical research and for the management of trimethylaminuria. J Am Diet Assoc. 2004;104(12):1836-1845. (PubMed)
- Wang Z, Tang WH, Buffa JA, et al. Prognostic value of choline and betaine depends on intestinal microbiota-generated metabolite trimethylamine-N-oxide. Eur Heart J. 2014;35(14):904-910. (PubMed)
- Heianza Y, Ma W, Manson JE, Rexrode KM, Qi L. Gut microbiota metabolites and risk of major adverse cardiovascular disease events and death: a systematic review and meta-analysis of prospective studies. J Am Heart Assoc. 2017;6(7):e004947. (PubMed)
- Papandreou C, More M, Bellamine A. Trimethylamine N-oxide in relation to cardiometabolic health-cause or effect? Nutrients. 2020;12(5):1330. (PubMed)
- Zeisel SH, Warrier M. Trimethylamine N-oxide, the microbiome, and heart and kidney disease. Annu Rev Nutr. 2017;37:157-181. (PubMed)
- Gatarek P, Kaluzna-Czaplinska J. Trimethylamine N-oxide (TMAO) in human health. EXCLI J. 2021;20:301-319. (PubMed)
- Canyelles M, Borras C, Rotllan N, Tondo M, Escola-Gil JC, Blanco-Vaca F. Gut microbiota-derived TMAO: a causal factor promoting atherosclerotic cardiovascular disease? Int J Mol Sci. 2023;24(3):1940. (PubMed)
- Al-Obaide MAI, Singh R, Datta P, et al. Gut microbiota-dependent trimethylamine-N-oxide and serum biomarkers in patients with T2DM and advanced CKD. J Clin Med. 2017;6(9):86. (PubMed)
- Zuo H, Svingen GFT, Tell GS, et al. Plasma concentrations and dietary intakes of choline and betaine in association with atrial fibrillation risk: results from 3 prospective cohorts with different health profiles. J Am Heart Assoc. 2018;7(8):e008190. (PubMed)
- Diez-Ricote L, San-Cristobal R, Concejo MJ, et al. One-year longitudinal association between changes in dietary choline or betaine intake and cardiometabolic variables in the PREvencion con DIeta MEDiterranea-Plus (PREDIMED-Plus) trial. Am J Clin Nutr. 2022;116(6):1565-1579. (PubMed)
- Hardwick RN, Clarke JD, Lake AD, et al. Increased susceptibility to methotrexate-induced toxicity in nonalcoholic steatohepatitis. Toxicol Sci. 2014;142(1):45-55. (PubMed)
- Lever M, George PM, Slow S, et al. Fibrates may cause an abnormal urinary betaine loss which is associated with elevations in plasma homocysteine. Cardiovasc Drugs Ther. 2009;23(5):395-401. (PubMed)
- Lever M, McEntyre CJ, George PM, et al. Extreme urinary betaine losses in type 2 diabetes combined with bezafibrate treatment are associated with losses of dimethylglycine and choline but not with increased losses of other osmolytes. Cardiovasc Drugs Ther. 2014;28(5):459-468. (PubMed)
- Lever M, George PM, Slow S, et al. Fibrates plus betaine: a winning combination? N Z Med J. 2010;123(1324):74-78. (PubMed)
Exclusión de Responsabilidades
El Centro de Informaciones sobre Micronutrientes del Instituto Linus Pauling entrega información científica relacionado con aspectos de importancia en salud pública de factores dietéticos, suplementos, comestibles y bebidas para el público en general. Esta información se entrega con el entendimiento que ni el autor ni la imprenta están entregando consejos de naturaleza médica, psicológica o nutricional. La información no debe ser usada para reemplazar la consulta con profesionales competentes en el área de cuidado de salud o de nutrición.
La información entregada con relación a factores y suplementos dietéticos, comestibles, y bebestibles, contenida en este sitio web, no cubre todos los usos, acciones, precauciones, efectos secundarios, e interacciones posibles. No debe ser considerado como consejo nutricional o médico para resolver problemas individuales. La responsabilidad legal por las acciones individuales u omisiones que se derivan del uso de este sitio está expresamente excluida.
No se puede copiar, modificar, distribuir, transmitir, realizar, publicar o vender ninguno de los materiales sujetos a derechos de autoría de este sitio web. Puede enlazar a este sitio web, pero debe incluir la siguiente declaración:
"Este enlace lleva a un sitio web proporcionado por el Instituto Linus Pauling en la Universidad Estatal de Oregon. [Su nombre] no está afiliado ni respaldado por el Instituto Linus Pauling o la Universidad Estatal de Oregon."