Contents
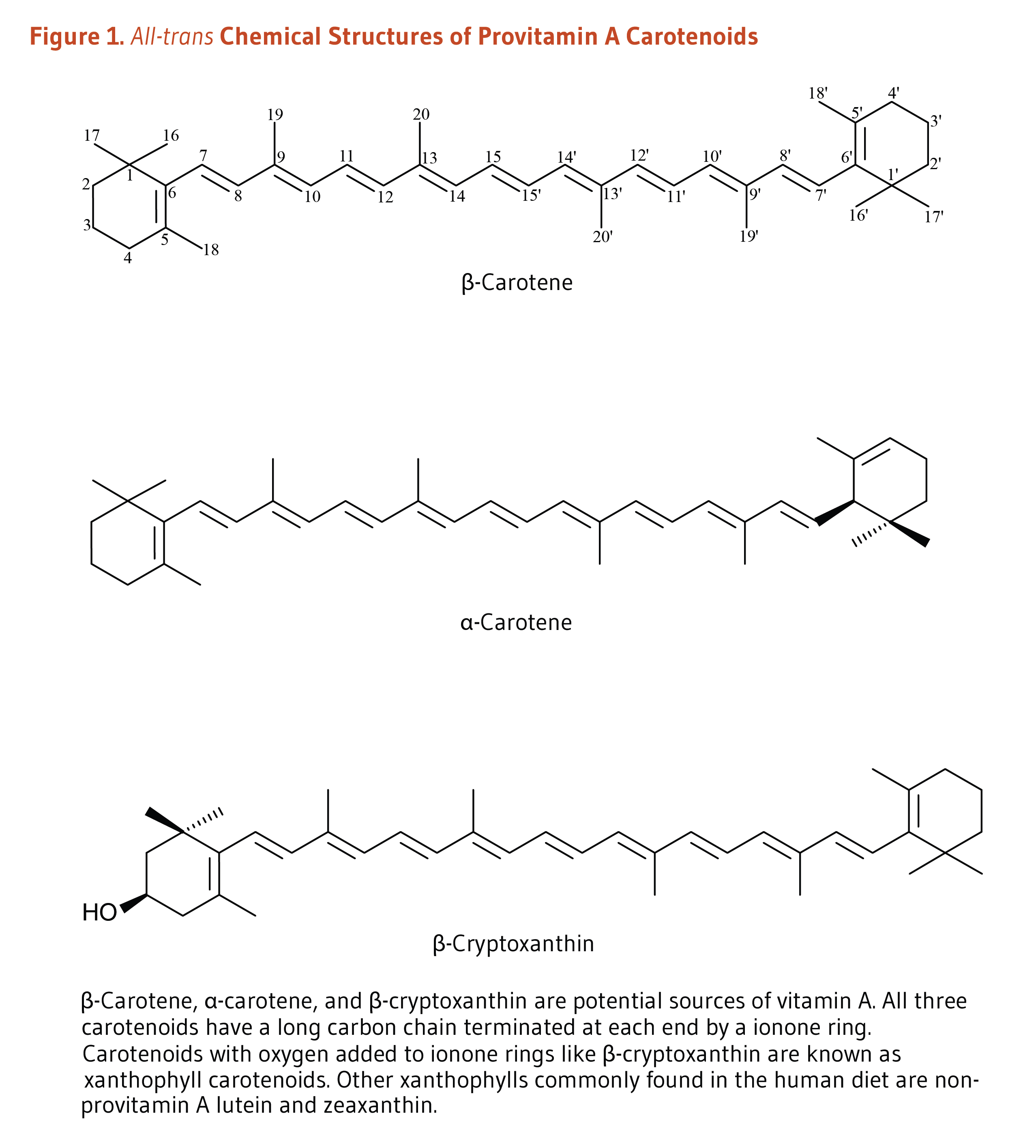
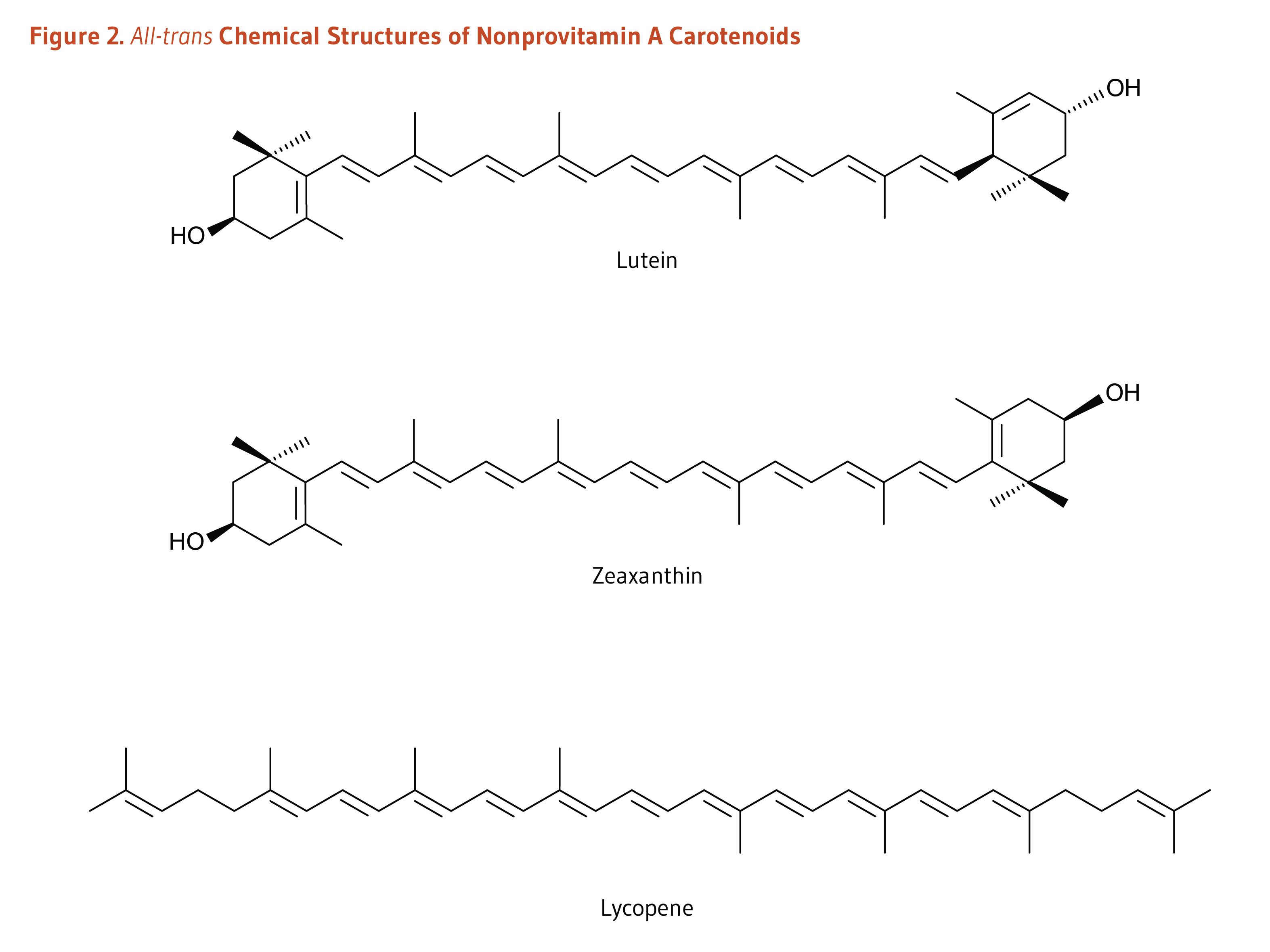
Carotenoids are a class of more than 750 naturally occurring pigments synthesized by plants, algae, and photosynthetic bacteria (1). These richly colored molecules are the sources of the yellow, orange, and red colors of many plants. Fruit and vegetables provide most of the 40 to 50 carotenoids found in the human diet. α-Carotene, β-carotene, β-cryptoxanthin, lutein, zeaxanthin, and lycopene are the most common dietary carotenoids (1). α-Carotene, β-carotene and β-cryptoxanthin are provitamin A carotenoids, meaning they can be converted by the body to retinol (Figure 1). Lutein, zeaxanthin, and lycopene are nonprovitamin A carotenoids because they cannot be converted to retinol (Figure 2).
For dietary carotenoids to be absorbed intestinally, they must be released from the food matrix and incorporated into mixed micelles (mixtures of bile salts and several types of lipids). Food processing and cooking help release carotenoids embedded in their food matrix and increase intestinal absorption (1). Moreover, carotenoid absorption requires the presence of fat in a meal. As little as 3 to 5 g of fat in a meal appears sufficient to ensure carotenoid absorption (2, 3), although the minimum amount of dietary fat required may be different for each carotenoid. The type of fat (e.g., medium-chain vs. long-chain triglycerides), the presence of soluble fiber, and the type and amount of carotenoids (e.g., esterified vs. non-esterified) in the food also appear to influence the rate and extent of carotenoid absorption (reviewed in 4). Because they do not need to be released from the plant matrix, carotenoid supplements (in oil) are more efficiently absorbed than carotenoids in food (3, 5). Although carotenoids were initially thought to be absorbed within the cells that line the intestine (enterocytes) only by passive diffusion, carotenoids are also actively absorbed via the apical membrane transporters, Scavenger Receptor-class B type I (SR-BI), Cluster Determinant 36 (CD36), and Niemann-Pick C1 like intracellular transporter 1 (NPC1L1) (6-8).
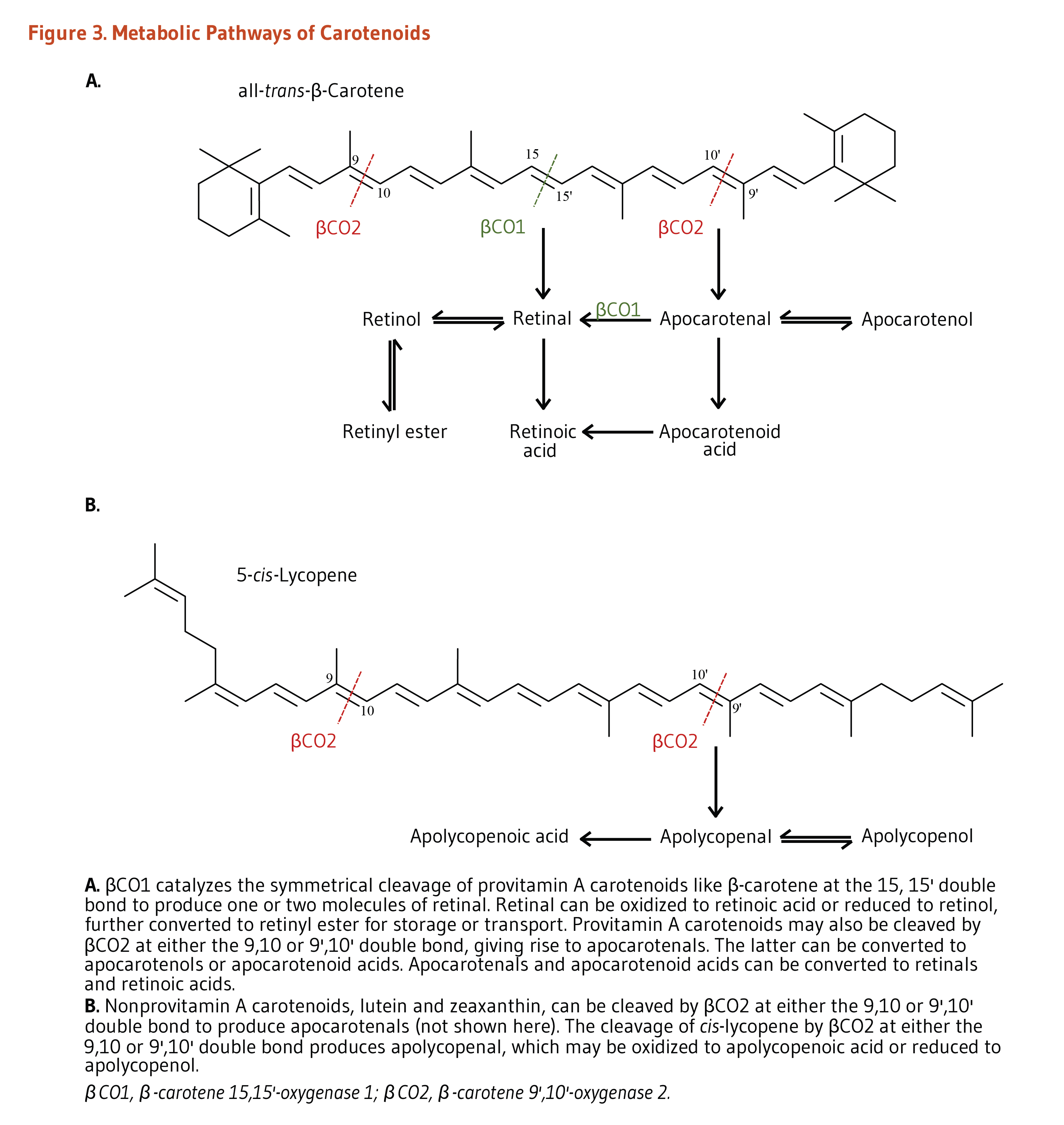
Within the enterocytes, provitamin A carotenoids may be cleaved by either β-carotene 15,15’-oxygenase 1 (BCO1) or by β-carotene 9’,10’-oxygenase 2 (BCO2) (Figure 3). BCO1 catalyzes the cleavage of provitamin A carotenoids into retinal, which is further reduced to retinol (vitamin A) or oxidized to retinoic acid (the biologically active form of vitamin A). β-Apocarotenal derived from the cleavage of β-carotene by BCO2 can be cleaved further by BCO1 to produce retinal. Although provitamin A carotenoids can be converted into apocarotenals by BCO2, the activity of this enzyme is higher toward nonprovitamin A carotenoids. Conversely, BCO1 shows limited affinity toward nonprovitamin A carotenoids (1).
Within the enterocytes, uncleaved carotenoids and retinyl esters (derived from retinol) are incorporated into triglyceride-rich lipoproteins called chylomicrons, secreted into lymphatic vessels, and then released into the bloodstream (1). Triglycerides are depleted from circulating chylomicrons through the activity of an enzyme called lipoprotein lipase, resulting in the formation of chylomicron remnants. Chylomicron remnants are taken up by the liver, where carotenoids can be cleaved by BCO1/BCO2 or incorporated into lipoproteins and secreted back into the circulation for delivery to extrahepatic tissues. Of note, more hydrophilic molecules in the enterocytes like retinoic acid and apocarotenals can be transported directly to the liver through the portal blood system.
The conversion of provitamin A carotenoids to retinol is influenced by the vitamin A status of the individual (9). The regulatory mechanism involving the intestine-specific homeobox (ISX) transcription factor can block carotenoid uptake and vitamin A production by inhibiting the expression of SR-BI and BCO1. ISX is under the control of retinoic acid and retinoic acid receptor (RAR)-dependent mechanisms such that, when vitamin A stores are high, ISX is activated and both provitamin A carotenoid absorption and conversion to retinol are inhibited. Conversely, during vitamin A insufficiency, the expression of both SR-BI and BCO1 is no longer repressed by ISX, allowing for provitamin A carotenoid absorption and conversion to retinol (1).
Interindividual variations in blood and tissue concentrations of carotenoids have been attributed to genetic differences among individuals. Specifically, a number of single nucleotide polymorphisms (SNPs) — corresponding to changes of one nucleotide in the sequence of genes — have been identified in genes coding for proteins involved in intestinal uptake, transport, and metabolism of carotenoids (10). Specifically, SNPs within genes coding for SR-BI, CD36, and BCO1 are suspected to affect the expression and/or activity of these proteins and, in turn, individual carotenoid status (10). For more information genetic variants affecting carotenoid status, see the review by Moran et al. (5).
Vitamin A is essential for normal growth and development, immune system function, and vision (see the article on Vitamin A). Currently, the only essential function of carotenoids recognized in humans is that of the provitamin A carotenoids, α-carotene, β-carotene, and β-cryptoxanthin, to serve as a source of vitamin A (11).
Provitamin A carotenoids are less easily absorbed than preformed vitamin A and must be converted to retinol and other retinoids by the body (see Figure 3). The efficiency of conversion of provitamin A carotenoids into retinol is highly variable, depending on factors like food matrix, food preparation, and one’s digestive and absorptive capacities (12).
The most recent international standard of measure for vitamin A is retinol activity equivalent (RAE), which represents vitamin A activity as retinol. It has been determined that 2 micrograms (µg) of β-carotene in oil provided as a supplement could be converted by the body to 1 µg of retinol, giving it an RAE ratio of 2:1. However, 12 µg of β-carotene from food are required to provide the body with 1 µg of retinol, giving dietary β-carotene an RAE ratio of 12:1. Other provitamin A carotenoids in food are less easily absorbed than β-carotene, resulting in RAE ratios of 24:1. RAE ratios are shown in Table 1.
In plants, carotenoids have the important antioxidant function of quenching (deactivating) singlet oxygen, an oxidant formed during photosynthesis (13). Test tube studies indicated that lycopene is one of the most effective quenchers of singlet oxygen among carotenoids (14). They also suggested that carotenoids could inhibit the oxidation of fats (i.e., lipid peroxidation) under certain conditions, but their actions in humans appear to be more complex (15). Although important for plants, the relevance of singlet oxygen quenching to human health is less clear (1).
Some evidence suggests that carotenoids and/or their metabolites may upregulate the expression of antioxidant and detoxifying enzymes via the activation of the nuclear factor erythroid 2-related factor 2 (Nrf2)-dependent pathway (reviewed in 16). Briefly, Nrf2 is a transcription factor that is bound to the protein Kelch-like ECH-associated protein 1 (Keap1) in the cytosol. Keap1 responds to oxidative stress signals by freeing Nrf2. Upon release, Nrf2 translocates to the nucleus and binds to the antioxidant response element (ARE) located in the promoter of genes coding for antioxidant/detoxifying enzymes and scavengers. Nrf2/ARE-dependent genes code for numerous mediators of the antioxidant response, including glutamate-cysteine ligase (GCL), glutathione S-transferases (GSTs), thioredoxin, NAD(P)H quinone oxidoreductase 1 (NQO-1), and heme oxygenase 1 (HO-1) (17). One study showed an increase in the level of the major antioxidant glutathione and a protection against TNFα-induced oxidative stress in retinal pigment epithelial cells (RPE) following lycopene-mediated Nrf2 activation and GCL induction (18). Nrf2 activation by lycopene also protected RPE against TNFα-mediated proinflammatory signaling involving nuclear factor-κB (NF-κB) activation and intercellular adhesion molecule-1 (ICAM-1) expression (18). Lycopene was shown to trigger Nrf2-mediated antioxidant pathway in various cell types (19-21). At present, evidence from animal and human studies is very limited (16).
The long system of alternating double and single bonds common to all carotenoids allows them to absorb light in the visible range of the spectrum (13). This feature has particular relevance to the eye, where lutein, zeaxanthin, and meso-zeaxanthin (derived from in vivo conversion from lutein) efficiently absorb blue light. Depending on the carotenoid pigment density at the center of the eye’s retina (macula), up to 90% of blue light can be absorbed by these pigments. Reducing the amount of short-wavelength light that reaches the critical visual structures of the eye may protect them from light-induced oxidative damage (22). Because the only source of these plant pigments in the eye is diet, a number of observational and intervention studies have examined the potential of dietary and supplemental lutein and zeaxanthin to protect against age-related eye diseases (see Age-related macular degeneration and Cataracts). Supplemental lutein, alone or with zeaxanthin, was found to improve contrast sensitivity and protect against visual fatigue in young and/or healthy individuals (23-26). Lutein has also been suggested to improve visual function through stimulating neuronal signaling efficiency in the eye (27).
Carotenoids can facilitate communication between neighboring cells grown in culture by stimulating the synthesis of connexin proteins (28). Connexins form pores (gap junctions) in cell membranes, allowing cells to communicate through the exchange of small molecules. This type of intercellular communication is important for maintaining cells in a differentiated state and is often lost in cancer cells. Carotenoids facilitate intercellular communication by increasing the expression of the gene encoding a connexin protein, an effect that appears unrelated to the vitamin A or antioxidant activities of various carotenoids (29) and involving a retinoic acid receptor (RAR)-independent mechanism (30).
Because vitamin A is essential for normal immune system function, it is difficult to determine whether the effects of provitamin A carotenoids are related to their vitamin A activity or other activities of carotenoids. Although some clinical trials have found that β-carotene supplementation improves several biomarkers of immune function (31-33), increasing intakes of lycopene and lutein — carotenoids without vitamin A activity — have not resulted in similar improvements in biomarkers of immune function (34-36).
Although consumption of provitamin A carotenoids (α-carotene, β-carotene, and β-cryptoxanthin) can prevent vitamin A deficiency (see the article on Vitamin A), no overt deficiency symptoms have been identified in people consuming low-carotenoid diets if they consume adequate vitamin A (11). After reviewing the published scientific research in 2000, the Food and Nutrition Board of the Institute of Medicine concluded that the existing evidence was insufficient to establish a recommended dietary allowance (RDA) or adequate intake (AI) for carotenoids. The Board has set an RDA for vitamin A (see the article on Vitamin A). Recommendations by the National Cancer Institute, American Cancer Society, and American Heart Association to consume a variety of fruit and vegetables daily are aimed, in part, at increasing intakes of carotenoids.
In the US, lung cancer is the leading cause of death by cancer among adults, representing about 20% of all cancer-related deaths (37).
Dietary carotenoids: Several large prospective cohort studies, including the Nurses’ Health Study (NHS) and the Health Professionals Follow-up Study (HPFS), have examined potential associations between carotenoid intake and/or blood concentrations and lung cancer (38). In a meta-analysis of eight prospective cohort studies, including NHS and HPFS, the highest versus lowest quantile of total carotenoid intake was significantly associated with a 21% reduced risk of lung cancer. For the individual carotenoids, the risk of lung cancer was estimated to be 20% and 14% lower with the highest versus lowest intakes of β-cryptoxanthin and lycopene, respectively. In contrast, dietary intakes of β-carotene, α-carotene, and lutein/zeaxanthin were not found to be significantly linked to a reduced risk of developing lung cancer (38). In addition, an analysis of the pooled results of 11 nested case-control and four prospective cohort studies found no association between serum concentrations of total carotenoids, β-carotene, α-lycopene, β-cryptoxanthin, and lutein/zeaxanthin and lung cancer. Only high versus low serum lycopene concentrations could be linked to a 29% lower risk of lung cancer (38). Any protective effect of dietary carotenoids against the development of lung cancer is likely small and not statistically significant (38).
Supplemental β-carotene: The effect of β-carotene supplementation on the risk of developing lung cancer has been examined in large randomized, placebo-controlled trials. In Finland, the Α-Tocopherol Βeta-Carotene (ATBC) cancer prevention trial evaluated the effects of 20 mg/day of β-carotene and/or 50 mg/day of α-tocopherol on more than 29,000 male smokers (39), and in the United States, the β-Carotene And Retinol Efficacy Trial (CARET) evaluated the effects of a combination of 30 mg/day of β-carotene and 25,000 IU/day of retinol (preformed vitamin A) in 18,314 men and women who were smokers, former smokers, or had a history of occupational asbestos exposure (40). Unexpectedly, the risk of lung cancer in the groups taking β-carotene supplements was increased by 16% after six years in the ATBC participants and by 28% after four years in the CARET participants. In contrast, the Physicians’ Health Study (PHS) examined the effect of β-carotene supplementation (50 mg every other day) on cancer risk in 22,071 male physicians in the United States, of whom only 11% were current smokers (41). In that lower risk population, β-carotene supplementation for more than 12 years was not associated with an increased risk of lung cancer. In addition, in the Linxian General Population trial conducted in about 29,000 Chinese participants, randomization to 15 mg/day of β-carotene, 30 mg/day of α-tocopherol, and 15 µg/day of selenium, was not found to be associated with lung cancer mortality 10 years after the intervention ended (42). Moreover, five-year follow-up of the Age-Related Eye Disease Study 2 (AREDS2) trial found that β-carotene supplementation nearly doubled the risk of developing lung cancer in former smokers compared to nonsmokers (current smokers did not receive β-carotene supplements) (43). AREDS2 was a multicenter, randomized double-blind, placebo-controlled trial that evaluated the effects of supplementation with antioxidant vitamins and minerals for five years to treat age-related macular degeneration (see below). Finally, a meta-analysis of four randomized controlled trials, including but not limited to trials in high-risk populations like smokers, found β-carotene supplementation (alone or with retinol; 3.7 to 12 years) increased the risk of lung cancer by 20% compared to control (OR, 1.20; 95% CI, 1.01-1.42) (44).
Although the reasons for the increase in lung cancer risk are not yet clear, several mechanisms have been proposed (45). Baseline β-carotene status might be one factor that influences whether with β-carotene supplementation promotes carcinogenesis in the lungs of smokers (46). The US Preventive Services Task Force estimated that the risks of high-dose β-carotene supplementation outweigh any potential benefits for cancer prevention and recommended against supplementation, especially in smokers or other high-risk populations (44, 47).
Prostate cancer is the one of the most prevalent cancers among US men, second only to non-melanoma skin cancer (48).
Dietary lycopene: Several early prospective cohort studies suggested that lycopene-rich diets were associated with significant reductions in the risk of prostate cancer, particularly more aggressive forms (49). Several pooled data analyses of observational studies that examining potential links between dietary intakes and/or circulating concentrations of lycopene and risk of prostate cancer have been completed. A 2015 meta-analysis of observational studies found no association of prostate cancer risk with dietary lycopene intakes (10 case-cohort and two prospective cohort studies) but an inverse association with blood lycopene concentrations (two case-control, nine nested case-control, and one cohort studies) (50, 51). Additionally, a meta-analysis of 15 nested case-control studies conducted by the Endogenous Hormones, Nutritional Biomarkers, and Prostate Cancer Collaborative Group showed an inverse association between circulating lycopene concentrations and risk of advanced stage and/or aggressive prostate cancer, while no association was found with risk of non-aggressive or localized disease (52). A 2017 meta-analysis of observational studies found inverse associations between both dietary (6 cohort/case-cohort and 15 case-control studies) and circulating (1 cohort study, 4 case-control studies, and 12 nested case-control studies) lycopene and prostate cancer risk; risk reductions were 12% in both analyses (RR, 0.88; 95% CI, 0.78-0.98) (53). However, this meta-analysis found no associations between dietary lycopene (5 studies) or circulating lycopene (6 studies) and advanced prostate cancer (53).
While there is considerable scientific interest in the potential for lycopene to help prevent prostate cancer, it is not yet clear whether the prostate cancer risk reduction observed in some observational studies is related to lycopene itself, other compounds in tomatoes, or other factors associated with lycopene-rich diets (54). Experimental studies in rodents suggest that lycopene is protective against prostate cancer but not the only protective compound found in tomatoes (reviewed in 5). Of note, the 2014 World Cancer Research Fund International report on Diet, Nutrition, Physical Activity, and Prostate Cancer suggested the need for better designed studies to establish whether consumption of lycopene-containing foods could be linked to a lower risk of prostate cancer (55).
Supplemental lycopene: To date, a few short-term, dietary intervention studies using lycopene in patients with precancerous prostate lesions (high grade prostatic intraepithelial neoplasia; HGPIN) or prostate cancer have been completed. Specifically, two small randomized controlled studies examined the effect of lycopene supplementation for up to six months in men with HGPIN (56, 57). The consumption of 30 to 35 mg/day of supplemental lycopene in the form of tomato extract (56), or together with selenium (55 mg/day) and green tea catechins (600 mg/day) (57), showed no benefit on the rate of progression to prostate cancer at six-month (56, 57) and 37-month follow-ups (57). Earlier small trials in men with HGPIN led to similar conclusions (reviewed in 58). Additionally, a randomized controlled trial in men with localized prostate cancer found that supplementation with 15, 30, or 45 mg of lycopene (until prostatectomy) did not significantly increase plasma lycopene concentration, modify the ratio of steroid hormones in blood, or reduce the concentration of markers of proliferation (i.e., prostate-specific antigen [PSA] and Ki-67) compared to placebo (59). In another trial, 54 patients with metastatic prostate cancer were randomized to orchidectomy alone or orchidectomy plus 4 mg/day of lycopene (60). The proportion of complete clinical response to treatment — assessed by serum PSA and/or bone scan returning to normal — and patient survival rate were found to be significantly higher in patients supplemented with lycopene (60).
Moreover, a meta-analysis of six randomized controlled trials in patients with non-metastatic prostate cancer found that supplemental lycopene (15-30 mg/day for 3 to 24 weeks) had no effect on circulating PSA concentrations (61). Yet, a subgroup analysis revealed a benefit of PSA reduction in patients with higher concentrations at baseline (PSA ≥6.5 µg/L) (61).
Large-scale, controlled clinical trials are needed to further examine the safety and efficacy of long-term use of lycopene supplements for prostate cancer prevention or treatment.
In a meta-analysis of 12 prospective cohort studies, no association was found between total and individual carotenoid intake and risk of breast cancer, except with β-carotene for which a 5% reduction in breast cancer risk was estimated for every 5 mg/day increment in consumption (62). In a pooled analysis of 14 nested case-control studies and one follow-up study of a clinical trial, reductions in breast cancer risk were found to be associated with blood concentrations of total carotenoids (-26%), α-carotene (-20%), and lutein (-30%) (62). Another study that recalibrated data for consistency across eight large prospective cohorts before pooled analysis found reduced breast cancer risk to be associated with the highest versus lowest quintile of blood concentrations of total carotenoids (-21%), β-carotene (-17%), and lycopene (-22%) (63). Further analyses found an inverse association between the blood concentrations of β-carotene and α-carotene and risk of estrogen receptor-negative (ER-), but not estrogen receptor-positive (ER+), breast tumors (63). A similar result was reported in a case-control study nested within the multicenter, large, European Prospective Investigation into Cancer and Nutrition (EPIC) study (64). In a nested case-control study of the Nurses’ Health Study (NHS) and NHSII, a 20% reduction in risk of breast cancer was seen in those with the highest total plasma carotenoids (≥142.1 µg/dL) compared to the lowest (<84.6 µg/dL), and this association was strongest in those presumed to be at higher risk of breast cancer (65). Protective associations were observed for higher circulating levels of the carotenoids, α-carotene (-20%), β-carotene (-18%), as well as lutein and zeaxanthin (-17%) (65). In contrast to the abovementioned pooled analysis (63), this study found a protective association between higher circulating carotenoids and ER+, but not ER-, breast tumors (65).
A case-control study nested within the EPIC study found a 31% lower risk of colorectal cancer with the highest versus lowest quartile of β-carotene intake, while no associations were shown with blood concentrations of other carotenoids or total carotenoids (66). The nested case-control study was included in a meta-analysis of 22 observational studies that failed to find associations between carotenoid intakes and colorectal cancer (67). A meta-analysis of 15 observational studies (11 case-control and 4 prospective cohort studies) reported no association between lycopene intake and colorectal cancer (68).
Pooled data also suggested that higher intakes of individual carotenoids, especially β-cryptoxanthin and lycopene, might be associated with a reduced risk of cancers of the mouth, pharynx, and larynx (69), but most of the data come from case-control control studies (70). The results of case-control studies are more likely to be distorted by bias than results of prospective cohort studies.
Examining blood concentrations of each carotenoid in relation to cancer subsites may help overcome limitations associated with dietary data and differences in carotenoid absorption. In a recent dose-response meta-analysis of observational studies, higher blood concentrations of several carotenoids, including α-carotene (3 studies), β-carotene (4 studies), as well as combined lutein and zeaxanthin (3 studies), were linked to a lower risk of developing bladder cancer (71).
In Western countries, degeneration of the macula, the center of the eye’s retina, is a leading cause of blindness among older adults. Long-term blue light exposure and oxidative damage in the outer segments of photoreceptors may lead to drusen and/or pigment abnormalities in the macula, increasing the risk of age-related macular degeneration (AMD) and central blindness.
Dietary lutein and zeaxanthin: The carotenoids found in the retina are lutein and zeaxanthin, which are both of dietary origin, and meso-zeaxanthin, which is derived in vivo conversion from lutein. These three carotenoids are present in high concentrations in the macula (known as macular pigment), where they are efficient absorbers of blue light. They may prevent a substantial amount of the blue light entering the eye from reaching the underlying structures involved in vision and protect against light-induced oxidative damage, which is thought to play a role in the pathology of age-related macular degeneration (reviewed in 22). It is also possible, though not proven, that lutein, zeaxanthin, and meso-zeaxanthin, act directly to neutralize oxidants formed in the retina.
Increasing dietary consumption of lutein and zeaxanthin was shown to raise their serum concentration and macular pigment density (72, 73). Some, but not all, observational studies have provided evidence that higher intakes of lutein and zeaxanthin are associated with lower risk of age-related macular degeneration (AMD) (74). While cross-sectional and retrospective case-control studies found that higher levels of lutein and zeaxanthin in the diet (75-77), blood (78, 79), and retina (80, 81) were associated with a lower incidence of AMD, several prospective cohort studies found no relationship between baseline dietary intakes or serum concentrations of lutein and zeaxanthin and the risk of developing AMD over time (82-85). One report examined the association between the incidence of AMD and calculated dietary intakes and predicted plasma concentrations of lutein and zeaxanthin in older adults (≥50 years) from two large prospective cohorts, the Nurses’ Health Study (NHS; 63,443 women) and the Health Professionals Follow-up Study (HPFS; 38,603 men), followed for 26 years and 24 years, respectively (86). The highest versus lowest quintile of predicted plasma lutein and zeaxanthin scores was associated with a 41% lower risk of advanced AMD, yet no association was found with intermediate AMD. Evidence has suggested that the consumption of about 6 mg/day of lutein and zeaxanthin from fruit and vegetables (compared with less than 2 mg/day) may decrease the risk of advanced AMD (77, 86). In a small population-based prospective study among 609 older adults in France, followed for a median of 7.6 years, found an inverse association between plasma lutein concentration at baseline and advanced AMD (HR, 0.63; 95% CI, 0.41-0.97) (87). No association was observed for circulating zeaxanthin and advanced AMD (87).
Supplemental lutein and zeaxanthin: The first randomized controlled trial in patients with atrophic AMD found that supplementation with 10 mg/day of lutein slightly improved visual acuity after one year compared to a placebo (88). The effects of long-term lutein supplementation on atrophic AMD were further investigated in combination with antioxidant vitamins and minerals in the Age-Related Eye Disease Study 2 (AREDS2), a multicenter, randomized, double-blind, placebo-controlled trial. In AREDS, oral supplementation with β-carotene (15 mg/day), vitamin C (500 mg/day), vitamin E (400 IU/day), zinc (80 mg/day as zinc oxide), and copper (2 mg/day as cupric oxide) for five years reduced the risk of developing advanced AMD by 25% (89). In the AREDS2 study conducted in 4,203 participants at risk for developing late AMD, supplemental lutein (10 mg/day) and zeaxanthin (2 mg/day), in combination with β-carotene, vitamin C, vitamin E, zinc, and copper (the ‘AREDS’ formulation), did not slow the progression to advanced AMD, although subgroup analyses revealed a benefit in those with the lowest dietary intakes of lutein + zeaxanthin (89). A total of 3,036 subjects were further randomized to various combinations of carotenoids; supplementation with lutein and zeaxanthin significantly reduced the risk of progression to late AMD and to neovascular AMD compared to supplementation with β-carotene (90).
Several smaller (n=30-433) randomized controlled trials also suggested that supplementation with xanthophyll carotenoids would be beneficial in the management of AMD (reviewed in 91). A meta-analysis of eight trials that examined the effect of supplemental lutein (6 to 20 mg/day) or/and zeaxanthin (0 to 10 mg/day) in 1,176 AMD subjects for up to 36 months found improvements in visual acuity and contrast sensitivity with increased levels of xanthophyll carotenoids (92). More recently, a randomized, placebo-controlled trial found daily consumption of a buttermilk drink with egg yolks enriched with lutein (1.4 mg), zeaxanthin (0.2 mg), and an omega-3 polyunsaturated fatty acid (DHA; 160 mg) for one year improved visual acuity and macular pigment optical density in subjects with drusen and/or retinal pigment abnormalities (about half of them being classified as having early AMD) (93). In a randomized, placebo-controlled trial in 74 patients with intermediate AMD, daily supplementation with lutein (10 mg/day) and zeaxanthin (2 mg) for two years, along with daily astaxanthin (4 mg), vitamin C (90 mg), vitamin E (30 mg), zinc (22.5 mg), copper (1 mg), and fish oil (containing 185 mg EPA and 140 mg DHA), resulted in a lower incidence of disease progression (2.1% of patients) when compared to placebo (15.4% of patients) (94).
Supplemental β-carotene: The first randomized controlled trial designed to examine the effect of a carotenoid supplement on AMD used β-carotene in combination with vitamin C, vitamin E, and zinc because lutein and zeaxanthin were not commercially available as supplements at the time the trial began (95). Although the combination of antioxidants and zinc lowered the risk of developing advanced macular degeneration in individuals with signs of moderate-to-severe macular degeneration in at least one eye, it is unlikely that the benefit was related to β-carotene since it is not present in the retina. Supplementation of male smokers in Finland with 20 mg/day of β-carotene for six years did not decrease the risk of AMD compared to placebo (96). A placebo-controlled trial in a cohort of 22,071 healthy US men found that β-carotene supplementation (50 mg every other day) had no effect on the incidence of age-related maculopathy — an early stage of AMD (97). Recent systematic reviews of randomized controlled trials have concluded that there is no evidence that β-carotene supplementation prevents or delays the onset of AMD (98, 99).
Retinopathy of prematurity
Preterm infants have an immature retinal vascular system that places them at risk of developing retinopathy of prematurity (ROP). Basically, preterm birth halts the normal development of retinal vascular system, which results in a retina that is poorly vascularized and highly susceptible to hyperoxia. In order to meet metabolic demand, the hypoxic retina induces the production of proangiogenic factors like VEGF (vascular endothelial growth factor). These factors stimulate the development of new blood vessels (angiogenesis), causing aberrant vessels sprouting from the retina into the vitreous. It is thought that an imbalance between the production of reactive oxygen species (ROS) and the reduced levels of antioxidants in preterm infants contributes to ROP pathogenesis and causes additional damage to the retina (reviewed in 100).
Supplemental lutein and zeaxanthin: A randomized controlled trial in 62 preterm infants (≤32 weeks of gestational age) failed to observe any benefits regarding ROP incidence and severity with lutein (0.5 mg/kg/day) and zeaxanthin (0.02 mg/kg/day) supplemented from the seventh day post birth until about 10 weeks of age (101). In two other multicenter, placebo-controlled trials in a total of 343 preterm infants, a daily oral dose of 0.14 mg of lutein and 0.006 mg of zeaxanthin administered from the first week after birth did not significantly reduce ROP incidence or the rate of progression from early to more advanced stages of ROP occurrence (102, 103). A fourth multicenter, randomized controlled trial in 203 preterm infants found that administration of a formula containing carotenoids (lutein/zeaxanthin, lycopene, and β-carotene) had no effect on ROP incidence but limited the progression to severe ROP stages in infants with mild ROP compared to a carotenoid-free formula (104). Compared with the control formula, preterm infants free of ROP fed the carotenoid-containing formula had significantly increased plasma carotenoid concentrations, which were correlated with greater rod photoreceptor sensitivity (104). More research is needed to examine whether carotenoid supplementation might promote normal photoreceptor development and prevent ROP in preterm infants.
Diabetic retinopathy
Supplementation with lutein and zeaxanthin has been shown to help preserve retinal integrity in diabetic rodents by reducing oxidative stress and inflammatory mediators (100). A higher ratio of plasma nonprovitamin A carotenoids (lutein, zeaxanthin, and lycopene) to vitamin A carotenoids (α-carotene, β-carotene, β-cryptoxanthin) was associated with a reduced risk of diabetic retinopathy in a cross-sectional analysis in 111 individuals with type 2 diabetes mellitus (105). A small observational study also found lower macular pigment optical density in those with type 2 diabetes (n=17) or mild nonproliferative diabetic retinopathy (n=12) compared to those without either condition (n=14) (106). Data from 1,430 individuals participating in the Atherosclerosis Risk In Communities (ARIC) study found no association between lutein intake and diabetic retinopathy after adjustment for confounding variables (107).
It is not known if supplementation with lutein and zeaxanthin might prevent or help treat diabetic retinopathy; well-designed, placebo-controlled studies would be needed to address these questions.
The primary function of the eye’s lens is to collect and focus light on the retina. Ultraviolet light and oxidants can damage proteins in the lens, causing structural changes that result in the formation of opacities known as cataracts. As people age, cumulative damage to lens proteins often results in cataracts that are large enough to interfere with vision (13). Risk factors other than aging and sunlight exposure include smoking, diabetes mellitus, higher body mass index, and use of estrogen replacement therapy. An estimated 50 million US adults are projected to be affected by age-related cataracts by 2050 (108).
Dietary lutein and zeaxanthin: The observation that lutein and zeaxanthin are the only carotenoids in the human lens has stimulated interest in the potential for increased intakes of lutein and zeaxanthin to prevent or slow the progression of cataracts (109). Large prospective cohort studies have found that men and women with the highest intakes of foods rich in lutein and zeaxanthin, particularly spinach, kale, and broccoli, were 18%-50% less likely to require cataract extraction (110, 111) or develop cataracts (112-114). Moreover, plasma concentrations of lutein and zeaxanthin have been found to be inversely associated to the progression of nuclear cataract. Additional research is required to determine whether these findings are related specifically to lutein and zeaxanthin intake or to other factors associated with diets high in carotenoid-rich foods (74).
Supplemental lutein and zeaxanthin: The Age-related Eye Disease Study 2 (AREDS2) failed to show an effect of supplemental lutein (10 mg/day) and zeaxanthin (2 mg/day; supplementation for a median of 4.7 years) on the risk of developing cataract, on the progression to severe cataract or to cataract surgery, and on visual acuity (115). However, the results might have been confounded by the fact that most participants were better nourished than the general population and/or used multivitamins that have been found to decrease the risk of developing cataract (116). Additional limitations to consider in interpreting the results have been reviewed elsewhere (115). Additional interventions are required to study whether supplemental lutein and zeaxanthin might be helpful in the prevention of cataract.
Supplemental β-carotene: Evidence from observational studies that cataracts were less prevalent in people with high dietary intakes and blood concentrations of carotenoids led to the inclusion of β-carotene supplements in several large randomized controlled trials of antioxidants. The results of those trials have been somewhat conflicting. β-Carotene supplementation (20 mg/day) for more than six years did not affect the prevalence of cataracts or the frequency of cataract surgery in male smokers living in Finland (96). In contrast, a 12-year study of male physicians in the US found that β-carotene supplementation (50 mg every other day) decreased the risk of cataracts in smokers but not in nonsmokers (117). Note that use of β-carotene supplements have been shown to increase the risk of lung cancer in smokers (see Supplemental β-carotene in lung cancer). Three randomized controlled trials examined the effect of an antioxidant combination that included β-carotene, vitamin C, and vitamin E on the progression of cataracts. Two trials found no benefit after supplementation for five years (118) or more than six years (119), but one trial found a small decrease in the progression of cataracts after three years of supplementation (120). Overall, the results of randomized controlled trials suggest that the benefit of β-carotene supplementation in slowing the progression of age-related cataracts does not outweigh the potential risks.
Dietary carotenoids: Because carotenoids are very soluble in fat and very insoluble in water, they circulate in lipoproteins, along with cholesterol and other fats. Evidence that low-density lipoprotein (LDL) oxidation plays a role in the development of atherosclerosis led scientists to investigate the role of antioxidant compounds like carotenoids in the prevention of cardiovascular disease (121). The thickness of the inner layers of the carotid arteries can be measured noninvasively using ultrasound technology. This measurement of carotid intima-media thickness is considered a reliable marker of atherosclerosis (122). A number of case-control and cross-sectional studies have found higher blood concentrations of carotenoids to be associated with significantly lower measures of carotid artery intima-media thickness (123-128). Additionally, higher plasma carotenoids at baseline have been associated with significant reductions in risk of cardiovascular disease in some prospective cohort studies (129-133) but not in others (134-137).
In a US national survey, National Health And Nutrition Examination Survey (NHANES) 2003-2006, serum total carotenoid concentration was inversely associated with blood concentrations of two cardiovascular risk factors, C-reactive protein (CRP) and total homocysteine (138). HDL-cholesterol concentration was found to be positively associated with α-carotene, β-cryptoxanthin, and lutein/zeaxanthin concentrations, but only the latter was inversely associated with LDL-cholesterol (138). NHANES 2007-2014 also found an inverse association between total dietary carotenoid intake (sum of α-carotene, β-carotene, β-cryptoxanthin, lycopene, lutein, and zeaxanthin) and risk of hypertension (139). Finally, a meta-analysis of observational studies reported lower risks of coronary heart disease (-12%) and stroke (-18%) in individuals in the highest versus lowest tertile of blood lutein concentration (140). In a recent meta-analysis, higher lycopene intake — from diet and/or supplements — was not associated with improvements in blood pressure or concentrations of blood lipids; the included studies were heterogeneous with respect to lycopene delivery (i.e., as a supplement or as food, varying tomato-containing products or extracts) and dose, characteristics of participants (healthy or with disease), and study duration (141).
While the results of several prospective studies indicate that people with higher intakes of carotenoid-rich fruit and vegetables are at lower risk of cardiovascular disease (137, 142-144), it is not yet clear whether this effect is a result of carotenoids or other factors associated with diets high in carotenoid-rich fruit and vegetables.
Supplemental β-carotene: In contrast to the results of observational studies suggesting that high dietary intakes of carotenoid-rich fruit and vegetables may decrease cardiovascular disease risk, four randomized controlled trials found no evidence that β-carotene supplements in doses ranging from 20 to 50 mg/day were effective in preventing cardiovascular disease (39, 41, 145, 146). Based on the results of these randomized controlled trials, the US Preventive Health Services Task Force found good evidence to suggest that β-carotene supplements provided no benefit in the prevention of cardiovascular disease in healthy adults (47). Moreover, a recent meta-analysis of 12 randomized, placebo-controlled trials found that β-carotene supplementation increased mortality related to cardiovascular disease (RR, 1.12; 95% CI, 1.04-1.19) (147). Thus, although diets rich in β-carotene have generally been associated with reduced cardiovascular disease risk in observational studies, there is no evidence that β-carotene supplementation reduces cardiovascular disease risk (148).
Supplemental lycopene: Several randomized controlled trials have examined whether supplementation with lycopene, tomato products, or tomato extracts might benefit cardiovascular health by improving blood pressure, lipid profiles, or function of the vascular endothelium. A recent meta-analysis of eight trials found no effect of supplemental lycopene on systolic or diastolic blood pressure; no benefits were found in healthy subjects or in hypertensive subjects (141). The meta-analysis also showed no effect of lycopene supplementation on total cholesterol (10 trials), LDL cholesterol (11 trials), or HDL (11 trials) cholesterol (141). Moreover, some, but not all (149, 150) short-term trials have indicated supplemental lycopene might improve function of the vascular endothelium in healthy subjects (151, 152); however, large, long-term placebo-controlled studies are needed.
An experimental study found that the administration of β-cryptoxanthin to ovariectomized mice limited bone resorption by inhibiting osteoclast differentiation but had no effect on osteoblast-driven bone formation (153). Yet, evidence of a protective role of β-cryptoxanthin — and other individual carotenoids — against bone loss in humans is scarce. In the prospective Framingham Osteoporosis Study, data analysis of 603 participants found no association between β-cryptoxanthin intake and changes in bone mineral density (BMD) over a four-year period (154). In contrast, intakes of total and individual carotenoids (β-carotene, lycopene, and lutein/zeaxanthin) were positively associated with proximal femur BMD in men over a four-year period, while lycopene intake was positively linked to lumbar spine BMD in women (154). The 17-year follow-up of participants in the Framingham Osteoporosis Study (946 participants) showed those in the highest tertile of total carotenoid intake (median intake: 23.7 mg/day) had a 51% lower risk of hip fracture compared to those in the lowest tertile (median intake: 7.3 mg/day) (155). In a much larger prospective study — the Singapore Chinese Health Study — in 63,257 men and women followed for 10 years, the highest versus lowest quartile of total carotenoid intake was associated with a 37% lower risk of hip fracture in men, but no association was observed in women (156). Dietary intakes of α- and β-carotene — but not of β-cryptoxanthin, lycopene, and lutein/zeaxanthin — were inversely associated with hip fracture risk in men (156). Nevertheless, in a small, four-year, Japanese prospective cohort study in 457 adults, the risk of osteoporosis was 93% lower in postmenopausal women in the highest versus lowest tertile of serum β-cryptoxanthin concentration (157). No such association was reported with serum concentrations of other individual carotenoids. More recently, in a cross-sectional analysis of the European Prospective Investigation into Cancer and Nutrition-Norfolk cohort, higher dietary intakes of β-carotene and combined lutein and zeaxanthin were linked to higher bone density at the heel in women (158). No associations of dietary carotenoid intake and heel bone density were found in men, and serum concentration of carotenoids was not linked to heel bone density in either men or women (158).
While a few studies have found a protective association between higher carotenoid intake and osteoporosis or bone fracture, the available studies are observational. Whether carotenoid supplementation may help prevent bone loss and reduce the risk of osteoporosis in older individuals is currently unknown; randomized controlled trials would be needed to address this question. It is important to note that high-dose supplementation with preformed vitamin A (retinol) has been associated with adverse effects on bone health (see the article on Vitamin A).
Observational studies have suggested that dietary lutein may be of benefit in maintaining cognitive health (159-162), and a cross-sectional study of 4,076 older adults associated higher blood lutein concentrations with improved cognition, including memory and executive function (163). As stated above, among the carotenoids, lutein and its isomer zeaxanthin are the only two that cross the blood-retina barrier to form macular pigment in the eye. Lutein also preferentially accumulates in the brain (164, 165). A few studies have suggested that lutein and zeaxanthin concentrations in the macula correlate with brain lutein and zeaxanthin status and therefore might be used as a biomarker of cognitive health (165-168). Additionally, in the Georgia Centenarian Study, the analysis of cross-sectional data from 47 centenarian decedents showed a positive association between post-mortem measures of brain lutein concentrations and pre-mortem measures of cognitive function (164). Brain lutein concentrations were found to be significantly lower in individuals with mild cognitive impairment compared to those with normal cognitive function (164). In a small, four-month, double-blind, placebo-controlled study in older women (ages, 60 to 80 years) without cognitive impairment, supplementation with lutein (12 mg/day) and zeaxanthin (~0.5 mg/day) significantly improved cognitive test performance (169). Two small trials in younger adults found supplementation with lutein (10 mg/day) and zeaxanthin (2 mg/day) for one year improved some measures of cognitive function, including memory (170, 171). However, the Age-related Eye Disease Study 2 (AREDS2) failed to show an effect of supplemental lutein (10 mg/day) and zeaxanthin (2 mg/day; supplementation for a median of 4.7 years) on the cognitive test scores of 3,741 older participants (mean age, 72.7 years) (172), perhaps because the trial was conducted in a highly educated and well-nourished population.
The most prevalent carotenoids in the human diet are α-carotene, β-carotene, β-cryptoxanthin, lycopene, lutein, and zeaxanthin (11). Most carotenoids in foods are found in the all-trans form (see Figure 1 and Figure 2 above), although cooking may result in the formation of other isomers. The relatively low bioavailability of carotenoids from most foods compared to supplements is partly due to the fact that they are associated with proteins in the plant matrix (173). Chopping, homogenizing, and cooking disrupt the plant matrix, increasing the bioavailability of carotenoids (3). For example, the bioavailability of lycopene from tomatoes is substantially improved by heating tomatoes in oil (174, 175). For information on other factors that affect carotenoid bioavailability, see above and the Moran et al. review (5).
α-Carotene and β-carotene are provitamin A carotenoids, meaning they can be converted in the body to vitamin A. The vitamin A activity of β-carotene in food is 1⁄12 that of retinol (preformed vitamin A). Thus, it would take 12 µg of β-carotene from food to provide the equivalent of 1 µg (0.001 mg) of retinol. The vitamin A activity of α-carotene from foods is 1⁄24 that of retinol, so it would take 24 µg of α-carotene from food to provide the equivalent of 1 µg of retinol. Orange and yellow vegetables like carrots and winter squash are rich sources of α- and β-carotene. Spinach is also a rich source of β-carotene, although the chlorophyll in spinach leaves hides the yellow-orange pigment. Some foods that are good sources of α-carotene and β-carotene are listed in Table 2 and Table 3 (176).
Like α- and β-carotene, β-cryptoxanthin is a provitamin A carotenoid. The vitamin A activity of β-cryptoxanthin from food was estimated to be 1⁄24 that of retinol, so it would take 24 µg of β-cryptoxanthin from food to provide the equivalent of 1 µg of retinol. Orange and red fruit and vegetables like sweet red peppers and oranges are particularly rich sources of β-cryptoxanthin. Some foods that are good sources of β-cryptoxanthin are listed in Table 4 (176).
Lycopene gives tomatoes, pink grapefruit, watermelon, and guava their red color. It has been estimated that 80% of the lycopene in the US diet comes from tomatoes and tomato products like tomato sauce, tomato paste, and ketchup (catsup) (177). Lycopene is not a provitamin A carotenoid because it cannot be converted to retinol. Some foods that are good sources of lycopene are listed in Table 5 (176).
Although lutein and zeaxanthin are different compounds, they are both classified as xanthophylls and nonprovitamin A carotenoids (see Figure 2 above). Some methods used to quantify lutein and zeaxanthin in food do not separate the two compounds, so they are typically reported as lutein and zeaxanthin or lutein + zeaxanthin. Both pigments are present in a variety of fruit and vegetables. Dark green leafy vegetables like spinach and kale are particularly rich sources of lutein but poor sources of zeaxanthin (178). Although relatively low in lutein, egg yolks and avocados are highly bioavailable sources of lutein. Good sources of dietary zeaxanthin include yellow corn, corn-based products, bell peppers, and egg yolk (179). Some foods containing lutein and zeaxanthin are listed in Table 6 (176).
For more information on the carotenoid content of certain foods, search USDA's FoodData Central database.
Dietary supplements providing purified carotenoids and combinations of carotenoids are commercially available in the US without a prescription. Carotenoids are best absorbed when taken with a meal containing fat.
Supplements containing a mixture of carotenoids may include α-carotene. As a provitamin A carotenoid, supplemental α-carotene can contribute to fulfill vitamin A requirements. It is not known whether the relative bioavailability of supplemental α-carotene is greater than that of dietary α-carotene.
Supplements containing a mixture of carotenoids may include α-cryptoxanthin. As a provitamin A carotenoid, supplemental α-cryptoxanthin can contribute to fulfill vitamin A requirements. It is not known whether the relative bioavailability of supplemental α-cryptoxanthin is greater than that of dietary α-cryptoxanthin.
β-Carotene is sold as individual supplements and also found in supplements marketed to promote visual health (180). Commercially available β-carotene supplements usually contain between 1.5 mg and 15 mg of either synthetic β-carotene or natural β-carotene (mainly from the algae Dunaliella salina) per softgel capsule (178). As a provitamin A carotenoid, β-carotene may be used to provide all or part of the vitamin A in multivitamin supplements. The provitamin A activity of β-carotene from supplements is much higher than that of β-carotene from food: it takes only 2 micrograms [µg] (0.002 mg) of β-carotene from supplements to provide 1 µg of retinol (preformed vitamin A) compared to 12 µg of dietary β-carotene.
Of note, the β-carotene content of supplements is often listed in international units (IU) rather than µg: 3,000 µg (3 mg) of supplemental β-carotene provides 5,000 IU of vitamin A.
Lycopene has no provitamin A activity. Synthetic lycopene and lycopene from natural sources, mainly tomatoes, are available as nutritional supplements containing up to 15 mg of lycopene per softgel capsule (178).
Lutein and zeaxanthin are not provitamin A carotenoids. Lutein and zeaxanthin supplements are available as free carotenoids (non-esterified) or as esters (esterified to fatty acids). Both forms appear to have comparable bioavailability (181). Many commercially available lutein and zeaxanthin supplements have much higher amounts of lutein than zeaxanthin (178). Supplements containing only lutein or zeaxanthin are also available.
Although β-carotene can be converted to vitamin A, the conversion of β-carotene to vitamin A decreases when body stores of vitamin A are high (see Absorption, Metabolism, and Bioavailability). This may explain why high doses of β-carotene have never been found to cause vitamin A toxicity. High doses of β-carotene (up to 180 mg/day) have been used to treat erythropoietic protoporphyria, a photosensitivity disorder, without toxic side effects (11).
No toxicities have been reported (182).
Increased lung cancer risk: Two randomized controlled trials in smokers and former asbestos workers found that supplementation with 20 to 30 mg/day of β-carotene for 4 to 6 years was associated with significant 16%-28% increases in the risk of lung cancer compared to placebo (see Supplemental β-carotene in lung cancer). Although the reasons for these findings are not yet clear, the potential risk of lung cancer in smokers and other high-risk groups supplemented with high-dose β-carotene outweigh any possible benefits for chronic disease prevention (47). It should be noted that there is no evidence that β-carotene supplementation may harm nonsmokers.
Carotenodermia: High doses of β-carotene supplements (≥30 mg/day) and the consumption of large amounts of carotene-rich food have resulted in a yellow discoloration of the skin (xanthoderma) known as carotenodermia, also called carotenemia. Carotenodermia is not associated with any underlying health problems and resolves when supplementation with β-carotene is discontinued or dietary carotene intake is reduced.
Lycopenodermia: High intakes of lycopene-rich food or supplements may result in a deep orange discoloration of the skin known as lycopenodermia. Because lycopene is more intensely colored than the carotenes, lycopenodermia may occur at lower doses than carotenodermia (11).
Lutein and zeaxanthin
A risk assessment analysis of 11 human studies concluded that lutein is likely safe at intake levels below 20 mg/day (183). A more recent case report documented "foveal sparkles" (eye crystals) in an older woman with glaucoma taking 20 mg/day of lutein for eight years; the patient also had very high dietary lutein intake, and total daily intake of lutein was not known (184).
β-Cryptoxanthin
A double-blind, placebo controlled trial in 90 young healthy women found that β-cryptoxanthin supplementation up to 6 mg/day for eight weeks was well tolerated (185).
Unlike vitamin A, high doses of β-carotene taken by pregnant women have not been associated with increased risk of birth defects (11). However, the safety of high-dose β-carotene supplements in pregnancy and lactation has not been well studied. Although there is no reason to limit dietary β-carotene intake, pregnant and breast-feeding women should avoid consuming more than 3 mg/day (1,500 µg RAE/day; 5,000 IU/day) of β-carotene from supplements unless they prescribed under medical supervision.
The safety of carotenoid supplements other than β-carotene in pregnancy and lactation has not been established, so pregnant and breast-feeding women should obtain carotenoids from food rather than supplements. There is no reason to limit the consumption of carotenoid-rich fruit and vegetables during pregnancy (178).
The cholesterol-lowering agents, cholestyramine (Questran) and colestipol (Colestid), can reduce absorption of fat-soluble vitamins and carotenoids, as can mineral oil and Orlistat (Xenical), a drug used to treat obesity (178). Colchicine, a drug used to treat gout, can cause intestinal malabsorption. However, long-term use of 1 to 2 mg/day of colchicine did not affect serum β-carotene concentrations in one study (186). Increasing gastric pH through the use of proton-pump inhibitors (Omeprazole, Lansoprazole) may decrease the absorption of a single dose of a β-carotene supplement, but the effect is unlikely to be clinically significant (187).
A three-year randomized controlled trial in 160 patients with documented coronary heart disease (CHD) and low serum high density lipoprotein (HDL) concentrations found that a combination of simvastatin (Zocor) and niacin increased HDL2 levels, inhibited the progression of coronary artery stenosis, and decreased the frequency of cardiovascular events, including myocardial infarction and stroke (188). Surprisingly, when an antioxidant combination of 1,000 mg of vitamin C, 536 mg of RRR-α-tocopherol (vitamin E), 100 µg of selenium, and 25 mg of β-carotene daily was taken with the simvastatin-niacin combination, the protective effects were diminished. Since the antioxidants were taken together in this trial, the individual contribution of β-carotene cannot be determined. In contrast, a much larger randomized controlled trial of simvastatin and an antioxidant combination of 297 mg of RRR-α-tocopherol, 250 mg of vitamin C, and 20 mg of β-carotene daily in more than 20,000 men and women with CHD or diabetes mellitus found that the antioxidant combination did not diminish the cardioprotective effects of simvastatin therapy over a five-year period (189). These contradictory findings indicate that further research is needed on potential interactions between antioxidant supplements and cholesterol-lowering agents, such as niacin and statins.
In a controlled feeding study, consumption of 18 g/day of the fat substitute Olestra (sucrose polyester; Olean) resulted in a 27% decrease in serum carotenoid concentrations after three weeks (190). Studies in people before and after the introduction of Olestra-containing snacks to the marketplace found that total serum carotenoid concentrations decreased by 15% in those who reported consuming at least 2 g/day of Olestra (191). One study in adults found that those who consumed more than 4.4 g of Olestra weekly experienced a 9.7% decline in total serum carotenoids compared to those not consuming Olestra (192).
Some studies found that the regular use of plant sterol-containing spreads resulted in modest, 10%-20% decreases in the plasma concentrations of some carotenoids, particularly α-carotene, β-carotene, and lycopene (see the article on Phytosterols) (193, 194). However, advising people who use plant sterol- or stanol-containing margarines to consume an extra serving of carotenoid-rich fruit or vegetables daily prevented decreases in plasma carotenoid concentrations (195).
The relationships between alcohol consumption and carotenoid metabolism are not well understood. There is some evidence that regular alcohol consumption inhibits the conversion of β-carotene to retinol (196). Increases in lung cancer risk associated with high-dose β-carotene supplementation in two randomized controlled trials were enhanced in those with higher alcohol intakes (40, 197).
The results of metabolic studies suggested that high doses of β-carotene compete with lutein and lycopene for absorption when consumed at the same time (198-200). However, the consumption of high-dose β-carotene supplements did not adversely affect serum carotenoid concentrations in long-term clinical trials (201-204).
Originally written in 2004 by:
Jane Higdon, Ph.D.
Linus Pauling Institute
Updated in December 2005 by:
Jane Higdon, Ph.D.
Linus Pauling Institute
Oregon State University
Updated in May 2009 by:
Victoria J. Drake, Ph.D.
Linus Pauling Institute
Oregon State University
Updated in July 2016 by:
Barbara Delage, Ph.D.
Linus Pauling Institute
Oregon State University
Updated in September 2023 by:
Victoria J. Drake, Ph.D.
Linus Pauling Institute
Oregon State University
Reviewed in October 2023 by:
Elizabeth J. Johnson, Ph.D., F.A.C.N., F.I.C.S.
Associate Professor,
Friedman School of Nutrition and Science & Policy
Tufts University
Copyright 2004-2024 Linus Pauling Institute
1. Wang XD. Carotenoids. In: Ross CA, Caballero B, Cousins RJ, Tucker KL, Ziegler TR, eds. Modern Nutrition in Health and Disease. 11th ed: Lippincott Williams & Wilkins; 2014:427-439.
2. Jalal F, Nesheim MC, Agus Z, Sanjur D, Habicht JP. Serum retinol concentrations in children are affected by food sources of beta-carotene, fat intake, and anthelmintic drug treatment. Am J Clin Nutr. 1998;68(3):623-629. (PubMed)
3. van Het Hof KH, West CE, Weststrate JA, Hautvast JG. Dietary factors that affect the bioavailability of carotenoids. J Nutr. 2000;130(3):503-506. (PubMed)
4. Priyadarshani AM. A review on factors influencing bioaccessibility and bioefficacy of carotenoids. Crit Rev Food Sci Nutr. 2015;57(8):1710-1717. (PubMed)
5. Moran NE, Mohn ES, Hason N, Erdman JW, Jr., Johnson EJ. Intrinsic and extrinsic factors impacting absorption, metabolism, and health effects of dietary carotenoids. Adv Nutr. 2018;9(4):465-492. (PubMed)
6. Reboul E. Absorption of vitamin A and carotenoids by the enterocyte: focus on transport proteins. Nutrients. 2013;5(9):3563-3581. (PubMed)
7. Bohn T, Desmarchelier C, Dragsted LO, et al. Host-related factors explaining interindividual variability of carotenoid bioavailability and tissue concentrations in humans. Mol Nutr Food Res. 2017;61(6):1600685. (PubMed)
8. Reboul E. Mechanisms of carotenoid intestinal absorption: where do we stand? Nutrients. 2019;11(4):838. (PubMed)
9. Tanumihardjo SA, Palacios N, Pixley KV. Provitamin a carotenoid bioavailability:what really matters? Int J Vitam Nutr Res. 2010;80(4-5):336-350. (PubMed)
10. Borel P. Genetic variations involved in interindividual variability in carotenoid status. Mol Nutr Food Res. 2012;56(2):228-240. (PubMed)
11. Food and Nutrition Board, Institute of Medicine. Beta-carotene and other carotenoids. Dietary reference intakes for vitamin C, vitamin E, selenium, and carotenoids. Washington, D.C.: National Academy Press; 2000:325-400. (National Academy Press)
12. Weber D, Grune T. The contribution of beta-carotene to vitamin A supply of humans. Mol Nutr Food Res. 2012;56(2):251-258. (PubMed)
13. Halliwell B, Gutteridge JMC. Free Radicals in Biology and Medicine. 3rd ed. New York, NY: Oxford University Press; 1999.
14. Di Mascio P, Kaiser S, Sies H. Lycopene as the most efficient biological carotenoid singlet oxygen quencher. Arch Biochem Biophys. 1989;274(2):532-538. (PubMed)
15. Young AJ, Lowe GM. Antioxidant and prooxidant properties of carotenoids. Arch Biochem Biophys. 2001;385(1):20-27. (PubMed)
16. Kaulmann A, Bohn T. Carotenoids, inflammation, and oxidative stress--implications of cellular signaling pathways and relation to chronic disease prevention. Nutr Res. 2014;34(11):907-929. (PubMed)
17. Ben-Dor A, Steiner M, Gheber L, et al. Carotenoids activate the antioxidant response element transcription system. Mol Cancer Ther. 2005;4(1):177-186. (PubMed)
18. Yang PM, Wu ZZ, Zhang YQ, Wung BS. Lycopene inhibits ICAM-1 expression and NF-kappaB activation by Nrf2-regulated cell redox state in human retinal pigment epithelial cells. Life Sci. 2016;155:94-101. (PubMed)
19. Lian F, Wang XD. Enzymatic metabolites of lycopene induce Nrf2-mediated expression of phase II detoxifying/antioxidant enzymes in human bronchial epithelial cells. Int J Cancer. 2008;123(6):1262-1268. (PubMed)
20. Sung LC, Chao HH, Chen CH, et al. Lycopene inhibits cyclic strain-induced endothelin-1 expression through the suppression of reactive oxygen species generation and induction of heme oxygenase-1 in human umbilical vein endothelial cells. Clin Exp Pharmacol Physiol. 2015;42(6):632-639. (PubMed)
21. Yang CM, Huang SM, Liu CL, Hu ML. Apo-8'-lycopenal induces expression of HO-1 and NQO-1 via the ERK/p38-Nrf2-ARE pathway in human HepG2 cells. J Agric Food Chem. 2012;60(6):1576-1585. (PubMed)
22. Krinsky NI, Landrum JT, Bone RA. Biologic mechanisms of the protective role of lutein and zeaxanthin in the eye. Annu Rev Nutr. 2003;23:171-201. (PubMed)
23. Kvansakul J, Rodriguez-Carmona M, Edgar DF, et al. Supplementation with the carotenoids lutein or zeaxanthin improves human visual performance. Ophthalmic Physiol Opt. 2006;26(4):362-371. (PubMed)
24. Ma L, Lin XM, Zou ZY, Xu XR, Li Y, Xu R. A 12-week lutein supplementation improves visual function in Chinese people with long-term computer display light exposure. Br J Nutr. 2009;102(2):186-190. (PubMed)
25. Stringham JM, Hammond BR. Macular pigment and visual performance under glare conditions. Optom Vis Sci. 2008;85(2):82-88. (PubMed)
26. Yagi A, Fujimoto K, Michihiro K, Goh B, Tsi D, Nagai H. The effect of lutein supplementation on visual fatigue: a psychophysiological analysis. Appl Ergon. 2009;40(6):1047-1054. (PubMed)
27. Stringham JM, Hammond BR, Jr. Dietary lutein and zeaxanthin: possible effects on visual function. Nutr Rev. 2005;63(2):59-64. (PubMed)
28. Bertram JS. Carotenoids and gene regulation. Nutr Rev. 1999;57(6):182-191. (PubMed)
29. Stahl W, Nicolai S, Briviba K, et al. Biological activities of natural and synthetic carotenoids: induction of gap junctional communication and singlet oxygen quenching. Carcinogenesis. 1997;18(1):89-92. (PubMed)
30. Vine AL, Leung YM, Bertram JS. Transcriptional regulation of connexin 43 expression by retinoids and carotenoids: similarities and differences. Mol Carcinog. 2005;43(2):75-85. (PubMed)
31. van Poppel G, Spanhaak S, Ockhuizen T. Effect of beta-carotene on immunological indexes in healthy male smokers. Am J Clin Nutr. 1993;57(3):402-407. (PubMed)
32. Hughes DA, Wright AJ, Finglas PM, et al. The effect of beta-carotene supplementation on the immune function of blood monocytes from healthy male nonsmokers. J Lab Clin Med. 1997;129(3):309-317. (PubMed)
33. Santos MS, Gaziano JM, Leka LS, Beharka AA, Hennekens CH, Meydani SN. Beta-carotene-induced enhancement of natural killer cell activity in elderly men: an investigation of the role of cytokines. Am J Clin Nutr. 1998;68(1):164-170. (PubMed)
34. Hughes DA, Wright AJ, Finglas PM, et al. Effects of lycopene and lutein supplementation on the expression of functionally associated surface molecules on blood monocytes from healthy male nonsmokers. J Infect Dis. 2000;182 Suppl 1:S11-15. (PubMed)
35. Watzl B, Bub A, Blockhaus M, et al. Prolonged tomato juice consumption has no effect on cell-mediated immunity of well-nourished elderly men and women. J Nutr. 2000;130(7):1719-1723. (PubMed)
36. Corridan BM, O'Donoghue M, Hughes DA, Morrissey PA. Low-dose supplementation with lycopene or beta-carotene does not enhance cell-mediated immunity in healthy free-living elderly humans. Eur J Clin Nutr. 2001;55(8):627-635. (PubMed)
37. American Cancer Society. Key Statistics for Lung Cancer. Available at: https://www.cancer.org/cancer/types/lung-cancer/about/key-statistics.html. Accessed 9/27/2023.
38. Gallicchio L, Boyd K, Matanoski G, et al. Carotenoids and the risk of developing lung cancer: a systematic review. Am J Clin Nutr. 2008;88(2):372-383. (PubMed)
39. The Alpha-Tocopherol Beta Carotene Cancer Prevention Study Group. The effect of vitamin E and beta carotene on the incidence of lung cancer and other cancers in male smokers. N Engl J Med. 1994;330(15):1029-1035. (PubMed)
40. Omenn GS, Goodman GE, Thornquist MD, et al. Risk factors for lung cancer and for intervention effects in CARET, the Beta-Carotene and Retinol Efficacy Trial. J Natl Cancer Inst. 1996;88(21):1550-1559. (PubMed)
41. Hennekens CH, Buring JE, Manson JE, et al. Lack of effect of long-term supplementation with beta carotene on the incidence of malignant neoplasms and cardiovascular disease. N Engl J Med. 1996;334(18):1145-1149. (PubMed)
42. Kamangar F, Qiao YL, Yu B, et al. Lung cancer chemoprevention: a randomized, double-blind trial in Linxian, China. Cancer Epidemiol Biomarkers Prev. 2006;15(8):1562-1564. (PubMed)
43. Chew EY, Clemons TE, Agron E, et al. Long-term outcomes of adding lutein/zeaxanthin and omega-3 fatty acids to the AREDS supplements on age-related macular degeneration progression: AREDS2 report 28. JAMA Ophthalmol. 2022;140(7):692-698. (PubMed)
44. O'Connor EA, Evans CV, Ivlev I, et al. Vitamin and mineral supplements for the primary prevention of cardiovascular disease and cancer: updated evidence report and systematic review for the US Preventive Services Task Force. JAMA. 2022;327(23):2334-2347. (PubMed)
45. Veeramachaneni S, Wang XD. Carotenoids and lung cancer prevention. Front Biosci (Schol Ed). 2009;1:258-274. (PubMed)
46. Mayne ST, Ferrucci LM, Cartmel B. Lessons learned from randomized clinical trials of micronutrient supplementation for cancer prevention. Annu Rev Nutr. 2012;32:369-390. (PubMed)
47. Moyer VA, US Preventive Services Task Force. Vitamin, mineral, and multivitamin supplements for the primary prevention of cardiovascular disease and cancer: U.S. Preventive services Task Force recommendation statement. Ann Intern Med. 2014;160(8):558-564. (PubMed)
48. Centers for Disease Control and Prevention. Prostate Cancer Statistics. Available at: https://www.cdc.gov/cancer/prostate/statistics/index.htm. Accessed 9/27/2023.
49. Giovannucci E. A review of epidemiologic studies of tomatoes, lycopene, and prostate cancer. Exp Biol Med (Maywood). 2002;227(10):852-859. (PubMed)
50. Wang Y, Cui R, Xiao Y, Fang J, Xu Q. Effect of carotene and lycopene on the risk of prostate cancer: a systematic review and dose-response meta-analysis of observational studies. PLoS One. 2015;10(9):e0137427. (PubMed)
51. Wang Y, Cui R, Xiao Y, Fang J, Xu Q. Correction: effect of carotene and lycopene on the risk of prostate cancer: a systematic review and dose-response meta-analysis of observational studies. PLoS One. 2015;10(10):e0140415. (PubMed)
52. Key TJ, Appleby PN, Travis RC, et al. Carotenoids, retinol, tocopherols, and prostate cancer risk: pooled analysis of 15 studies. Am J Clin Nutr. 2015;102(5):1142-1157. (PubMed)
53. Rowles JL, 3rd, Ranard KM, Smith JW, An R, Erdman JW, Jr. Increased dietary and circulating lycopene are associated with reduced prostate cancer risk: a systematic review and meta-analysis. Prostate Cancer Prostatic Dis. 2017;20(4):361-377. (PubMed)
54. Melendez-Martinez AJ, Mapelli-Brahm P, Benitez-Gonzalez A, Stinco CM. A comprehensive review on the colorless carotenoids phytoene and phytofluene. Arch Biochem Biophys. 2015;572:188-200. (PubMed)
55. World Cancer Research Fund International/American Institute for Cancer Research Continuous Update Project Report. Diet, Nutrition, Physical Activity, and Prostate Cancer 2014.
56. Gann PH, Deaton RJ, Rueter EE, et al. A phase II randomized trial of lycopene-rich tomato extract among men with high-grade prostatic intraepithelial neoplasia. Nutr Cancer. 2015;67(7):1104-1112. (PubMed)
57. Gontero P, Marra G, Soria F, et al. A randomized double-blind placebo controlled phase I-II study on clinical and molecular effects of dietary supplements in men with precancerous prostatic lesions. Chemoprevention or "chemopromotion"? Prostate. 2015;75(11):1177-1186. (PubMed)
58. Holzapfel NP, Holzapfel BM, Champ S, Feldthusen J, Clements J, Hutmacher DW. The potential role of lycopene for the prevention and therapy of prostate cancer: from molecular mechanisms to clinical evidence. Int J Mol Sci. 2013;14(7):14620-14646. (PubMed)
59. Kumar NB, Besterman-Dahan K, Kang L, et al. Results of a randomized clinical trial of the action of several doses of lycopene in localized prostate cancer: administration prior to radical prostatectomy. Clin Med Urol. 2008;1:1-14. (PubMed)
60. Ansari MS, Gupta NP. A comparison of lycopene and orchidectomy vs orchidectomy alone in the management of advanced prostate cancer. BJU Int. 2003;92(4):375-378; discussion 378. (PubMed)
61. Sadeghian M, Asadi M, Rahmani S, Sadeghi N, Hosseini SA, Zare Javid A. Lycopene does not affect prostate-specific antigen in men with non-metastatic prostate cancer: a systematic review and meta-analysis of randomized controlled trials. Nutr Cancer. 2021;73(11-12):2796-2807. (PubMed)
62. Aune D, Chan DS, Vieira AR, et al. Dietary compared with blood concentrations of carotenoids and breast cancer risk: a systematic review and meta-analysis of prospective studies. Am J Clin Nutr. 2012;96(2):356-373. (PubMed)
63. Eliassen AH, Hendrickson SJ, Brinton LA, et al. Circulating carotenoids and risk of breast cancer: pooled analysis of eight prospective studies. J Natl Cancer Inst. 2012;104(24):1905-1916. (PubMed)
64. Bakker MF, Peeters PH, Klaasen VM, et al. Plasma carotenoids, vitamin C, tocopherols, and retinol and the risk of breast cancer in the European Prospective Investigation into Cancer and Nutrition cohort. Am J Clin Nutr. 2016;103(2):454-464. (PubMed)
65. Peng C, Gao C, Lu D, et al. Circulating carotenoids and breast cancer among high-risk individuals. Am J Clin Nutr. 2021;113(3):525-533. (PubMed)
66. Leenders M, Leufkens AM, Siersema PD, et al. Plasma and dietary carotenoids and vitamins A, C and E and risk of colon and rectal cancer in the European Prospective Investigation into Cancer and Nutrition. Int J Cancer. 2014;135(12):2930-2939. (PubMed)
67. Panic N, Nedovic D, Pastorino R, Boccia S, Leoncini E. Carotenoid intake from natural sources and colorectal cancer: a systematic review and meta-analysis of epidemiological studies. Eur J Cancer Prev. 2017;26(1):27-37. (PubMed)
68. Wang X, Yang HH, Liu Y, Zhou Q, Chen ZH. Lycopene consumption and risk of colorectal cancer: a meta-analysis of observational studies. Nutr Cancer. 2016;68(7):1083-1096. (PubMed)
69. Leoncini E, Nedovic D, Panic N, Pastorino R, Edefonti V, Boccia S. Carotenoid intake from natural sources and head and neck cancer: a systematic review and meta-analysis of epidemiological studies. Cancer Epidemiol Biomarkers Prev. 2015;24(7):1003-1011. (PubMed)
70. Brewczynski A, Jablonska B, Kentnowski M, Mrowiec S, Skladowski K, Rutkowski T. The association between carotenoids and head and neck cancer risk. Nutrients. 2021;14(1):88. (PubMed)
71. Wu S, Liu Y, Michalek JE, et al. Carotenoid intake and circulating carotenoids are inversely associated with the risk of bladder cancer: a dose-response meta-analysis. Adv Nutr. 2020;11(3):630-643. (PubMed)
72. Hammond BR, Jr., Johnson EJ, Russell RM, et al. Dietary modification of human macular pigment density. Invest Ophthalmol Vis Sci. 1997;38(9):1795-1801. (PubMed)
73. Mares JA, LaRowe TL, Snodderly DM, et al. Predictors of optical density of lutein and zeaxanthin in retinas of older women in the Carotenoids in Age-Related Eye Disease Study, an ancillary study of the Women's Health Initiative. Am J Clin Nutr. 2006;84(5):1107-1122. (PubMed)
74. Mares-Perlman JA, Millen AE, Ficek TL, Hankinson SE. The body of evidence to support a protective role for lutein and zeaxanthin in delaying chronic disease. Overview. J Nutr. 2002;132(3):518S-524S. (PubMed)
75. Snellen EL, Verbeek AL, Van Den Hoogen GW, Cruysberg JR, Hoyng CB. Neovascular age-related macular degeneration and its relationship to antioxidant intake. Acta Ophthalmol Scand. 2002;80(4):368-371. (PubMed)
76. Mares-Perlman JA, Fisher AI, Klein R, et al. Lutein and zeaxanthin in the diet and serum and their relation to age-related maculopathy in the third national health and nutrition examination survey. Am J Epidemiol. 2001;153(5):424-432. (PubMed)
77. Seddon JM, Ajani UA, Sperduto RD, et al. Dietary carotenoids, vitamins A, C, and E, and advanced age-related macular degeneration. Eye Disease Case-Control Study Group. JAMA. 1994;272(18):1413-1420. (PubMed)
78. Antioxidant status and neovascular age-related macular degeneration. Eye Disease Case-Control Study Group. Arch Ophthalmol. 1993;111(1):104-109. (PubMed)
79. Gale CR, Hall NF, Phillips DI, Martyn CN. Lutein and zeaxanthin status and risk of age-related macular degeneration. Invest Ophthalmol Vis Sci. 2003;44(6):2461-2465. (PubMed)
80. Bone RA, Landrum JT, Mayne ST, Gomez CM, Tibor SE, Twaroska EE. Macular pigment in donor eyes with and without AMD: a case-control study. Invest Ophthalmol Vis Sci. 2001;42(1):235-240. (PubMed)
81. Beatty S, Murray IJ, Henson DB, Carden D, Koh H, Boulton ME. Macular pigment and risk for age-related macular degeneration in subjects from a Northern European population. Invest Ophthalmol Vis Sci. 2001;42(2):439-446. (PubMed)
82. Cho E, Seddon JM, Rosner B, Willett WC, Hankinson SE. Prospective study of intake of fruits, vegetables, vitamins, and carotenoids and risk of age-related maculopathy. Arch Ophthalmol. 2004;122(6):883-892. (PubMed)
83. Flood V, Smith W, Wang JJ, Manzi F, Webb K, Mitchell P. Dietary antioxidant intake and incidence of early age-related maculopathy: the Blue Mountains Eye Study. Ophthalmology. 2002;109(12):2272-2278. (PubMed)
84. Mares-Perlman JA, Klein R, Klein BE, et al. Association of zinc and antioxidant nutrients with age-related maculopathy. Arch Ophthalmol. 1996;114(8):991-997. (PubMed)
85. Mares-Perlman JA, Brady WE, Klein R, et al. Serum antioxidants and age-related macular degeneration in a population-based case-control study. Arch Ophthalmol. 1995;113(12):1518-1523. (PubMed)
86. Wu J, Cho E, Willett WC, Sastry SM, Schaumberg DA. Intakes of lutein, zeaxanthin, and other carotenoids and age-related macular degeneration during 2 decades of prospective follow-up. JAMA Ophthalmol. 2015;133(12):1415-1424. (PubMed)
87. Merle BMJ, Cougnard-Gregoire A, Korobelnik JF, et al. Plasma lutein, a nutritional biomarker for development of advanced age-related macular degeneration: the Alienor study. Nutrients. 2021;13(6):2047. (PubMed)
88. Richer S, Stiles W, Statkute L, et al. Double-masked, placebo-controlled, randomized trial of lutein and antioxidant supplementation in the intervention of atrophic age-related macular degeneration: the Veterans LAST study (Lutein Antioxidant Supplementation Trial). Optometry. 2004;75(4):216-230. (PubMed)
89. Age-Related Eye Disease Study 2 Research Group. Lutein + zeaxanthin and omega-3 fatty acids for age-related macular degeneration: the Age-Related Eye Disease Study 2 (AREDS2) randomized clinical trial. JAMA. 2013;309(19):2005-2015. (PubMed)
90. Age-Related Eye Disease Study 2 Research Group, Chew EY, Clemons TE, et al. Secondary analyses of the effects of lutein/zeaxanthin on age-related macular degeneration progression: AREDS2 report No. 3. JAMA Ophthalmol. 2014;132(2):142-149. (PubMed)
91. Scripsema NK, Hu DN, Rosen RB. Lutein, zeaxanthin, and meso-zeaxanthin in the clinical management of eye disease. J Ophthalmol. 2015;2015:865179. (PubMed)
92. Liu R, Wang T, Zhang B, et al. Lutein and zeaxanthin supplementation and association with visual function in age-related macular degeneration. Invest Ophthalmol Vis Sci. 2015;56(1):252-258. (PubMed)
93. van der Made SM, Kelly ER, Kijlstra A, Plat J, Berendschot TT. Increased macular pigment optical density and visual acuity following consumption of a buttermilk drink containing lutein-enriched egg yolks: a randomized, double-blind, placebo-controlled trial. J Ophthalmol. 2016;2016:9035745. (PubMed)
94. Piatti A, Croce A, Mazzacane D, et al. Effect of 2-year nutritional supplementation on progression of age-related macular degeneration. Eur J Ophthalmol. 2020;30(2):376-381. (PubMed)
95. Age-Related Eye Disease Study Research Group. A randomized, placebo-controlled, clinical trial of high-dose supplementation with vitamins C and E, beta carotene, and zinc for age-related macular degeneration and vision loss: AREDS report no. 8. Arch Ophthalmol. 2001;119(10):1417-1436. (PubMed)
96. Teikari JM, Laatikainen L, Virtamo J, et al. Six-year supplementation with alpha-tocopherol and beta-carotene and age-related maculopathy. Acta Ophthalmol Scand. 1998;76(2):224-229. (PubMed)
97. Christen WG, Manson JE, Glynn RJ, et al. Beta carotene supplementation and age-related maculopathy in a randomized trial of US physicians. Arch Ophthalmol. 2007;125(3):333-339. (PubMed)
98. Evans J. Antioxidant supplements to prevent or slow down the progression of AMD: a systematic review and meta-analysis. Eye. 2008;22(6):751-760. (PubMed)
99. Evans JR, Henshaw K. Antioxidant vitamin and mineral supplements for preventing age-related macular degeneration. Cochrane Database Syst Rev. 2008(1):CD000253. (PubMed)
100. Gong X, Rubin LP. Role of macular xanthophylls in prevention of common neovascular retinopathies: retinopathy of prematurity and diabetic retinopathy. Arch Biochem Biophys. 2015;572:40-48. (PubMed)
101. Romagnoli C, Giannantonio C, Cota F, et al. A prospective, randomized, double blind study comparing lutein to placebo for reducing occurrence and severity of retinopathy of prematurity. J Matern Fetal Neonatal Med. 2011;24 Suppl 1:147-150. (PubMed)
102. Dani C, Lori I, Favelli F, et al. Lutein and zeaxanthin supplementation in preterm infants to prevent retinopathy of prematurity: a randomized controlled study. J Matern Fetal Neonatal Med. 2012;25(5):523-527. (PubMed)
103. Manzoni P, Guardione R, Bonetti P, et al. Lutein and zeaxanthin supplementation in preterm very low-birth-weight neonates in neonatal intensive care units: a multicenter randomized controlled trial. Am J Perinatol. 2013;30(1):25-32. (PubMed)
104. Rubin LP, Chan GM, Barrett-Reis BM, et al. Effect of carotenoid supplementation on plasma carotenoids, inflammation and visual development in preterm infants. J Perinatol. 2012;32(6):418-424. (PubMed)
105. Brazionis L, Rowley K, Itsiopoulos C, O'Dea K. Plasma carotenoids and diabetic retinopathy. Br J Nutr. 2009;101(2):270-277. (PubMed)
106. Lima VC, Rosen RB, Maia M, et al. Macular pigment optical density measured by dual-wavelength autofluorescence imaging in diabetic and nondiabetic patients: a comparative study. Invest Ophthalmol Vis Sci. 2010;51(11):5840-5845. (PubMed)
107. Sahli MW, Mares JA, Meyers KJ, et al. Dietary intake of lutein and diabetic retinopathy in the Atherosclerosis Risk in Communities Study (ARIC). Ophthalmic Epidemiol. 2016;23(2):99-108. (PubMed)
108. The National Institutes of Heath/National Eye Institute. Cataracts. Available at: https://nei.nih.gov/eyedata/cataract. Accessed 6/9/2016.
109. Yeum KJ, Shang FM, Schalch WM, Russell RM, Taylor A. Fat-soluble nutrient concentrations in different layers of human cataractous lens. Curr Eye Res. 1999;19(6):502-505. (PubMed)
110. Brown L, Rimm EB, Seddon JM, et al. A prospective study of carotenoid intake and risk of cataract extraction in US men. Am J Clin Nutr. 1999;70(4):517-524. (PubMed)
111. Chasan-Taber L, Willett WC, Seddon JM, et al. A prospective study of carotenoid and vitamin A intakes and risk of cataract extraction in US women. Am J Clin Nutr. 1999;70(4):509-516. (PubMed)
112. Lyle BJ, Mares-Perlman JA, Klein BE, Klein R, Greger JL. Antioxidant intake and risk of incident age-related nuclear cataracts in the Beaver Dam Eye Study. Am J Epidemiol. 1999;149(9):801-809. (PubMed)
113. Christen WG, Liu S, Glynn RJ, Gaziano JM, Buring JE. Dietary carotenoids, vitamins C and E, and risk of cataract in women: a prospective study. Arch Ophthalmol. 2008;126(1):102-109. (PubMed)
114. Moeller SM, Voland R, Tinker L, et al. Associations between age-related nuclear cataract and lutein and zeaxanthin in the diet and serum in the carotenoids in the Age-Related Eye Disease Study, an ancillary study of the Women's Health Initiative. Arch Ophthalmol. 2008;126(3):354-364. (PubMed)
115. Age-Related Eye Disease Study 2 Research Group, Chew EY, SanGiovanni JP, et al. Lutein/zeaxanthin for the treatment of age-related cataract: AREDS2 randomized trial report no. 4. JAMA Ophthalmol. 2013;131(7):843-850. (PubMed)
116. Glaser TS, Doss LE, Shih G, et al. The association of dietary lutein plus zeaxanthin and B vitamins with cataracts in the Age-Related Eye Disease Study: AREDS Report No. 37. Ophthalmology. 2015;122(7):1471-1479. (PubMed)
117. Christen WG, Manson JE, Glynn RJ, et al. A randomized trial of beta carotene and age-related cataract in US physicians. Arch Ophthalmol. 2003;121(3):372-378. (PubMed)
118. Gritz DC, Srinivasan M, Smith SD, et al. The Antioxidants in Prevention of Cataracts Study: effects of antioxidant supplements on cataract progression in South India. Br J Ophthalmol. 2006;90(7):847-851. (PubMed)
119. Age-Related Eye Disease Study Research Group. A randomized, placebo-controlled, clinical trial of high-dose supplementation with vitamins C and E and beta carotene for age-related cataract and vision loss: AREDS report no. 9. Arch Ophthalmol. 2001;119(10):1439-1452. (PubMed)
120. Chylack LT, Jr., Brown NP, Bron A, et al. The Roche European American Cataract Trial (REACT): a randomized clinical trial to investigate the efficacy of an oral antioxidant micronutrient mixture to slow progression of age-related cataract. Ophthalmic Epidemiol. 2002;9(1):49-80. (PubMed)
121. Kritchevsky SB. beta-Carotene, carotenoids and the prevention of coronary heart disease. J Nutr. 1999;129(1):5-8. (PubMed)
122. Bots ML, Grobbee DE. Intima media thickness as a surrogate marker for generalised atherosclerosis. Cardiovasc Drugs Ther. 2002;16(4):341-351. (PubMed)
123. Rissanen TH, Voutilainen S, Nyyssonen K, Salonen R, Kaplan GA, Salonen JT. Serum lycopene concentrations and carotid atherosclerosis: the Kuopio Ischaemic Heart Disease Risk Factor Study. Am J Clin Nutr. 2003;77(1):133-138. (PubMed)
124. Dwyer JH, Paul-Labrador MJ, Fan J, Shircore AM, Merz CN, Dwyer KM. Progression of carotid intima-media thickness and plasma antioxidants: the Los Angeles Atherosclerosis Study. Arterioscler Thromb Vasc Biol. 2004;24(2):313-9. (PubMed)
125. McQuillan BM, Hung J, Beilby JP, Nidorf M, Thompson PL. Antioxidant vitamins and the risk of carotid atherosclerosis. The Perth Carotid Ultrasound Disease Assessment study (CUDAS). J Am Coll Cardiol. 2001;38(7):1788-1794. (PubMed)
126. Rissanen T, Voutilainen S, Nyyssonen K, Salonen R, Salonen JT. Low plasma lycopene concentration is associated with increased intima-media thickness of the carotid artery wall. Arterioscler Thromb Vasc Biol. 2000;20(12):2677-2681. (PubMed)
127. D'Odorico A, Martines D, Kiechl S, et al. High plasma levels of alpha- and beta-carotene are associated with a lower risk of atherosclerosis: results from the Bruneck study. Atherosclerosis. 2000;153(1):231-239. (PubMed)
128. Iribarren C, Folsom AR, Jacobs DR, Jr., Gross MD, Belcher JD, Eckfeldt JH. Association of serum vitamin levels, LDL susceptibility to oxidation, and autoantibodies against MDA-LDL with carotid atherosclerosis. A case-control study. The ARIC Study Investigators. Atherosclerosis Risk in Communities. Arterioscler Thromb Vasc Biol. 1997;17(6):1171-1177. (PubMed)
129. Sesso HD, Buring JE, Norkus EP, Gaziano JM. Plasma lycopene, other carotenoids, and retinol and the risk of cardiovascular disease in women. Am J Clin Nutr. 2004;79(1):47-53. (PubMed)
130. Rissanen TH, Voutilainen S, Nyyssonen K, et al. Low serum lycopene concentration is associated with an excess incidence of acute coronary events and stroke: the Kuopio Ischaemic Heart Disease Risk Factor Study. Br J Nutr. 2001;85(6):749-754. (PubMed)
131. Street DA, Comstock GW, Salkeld RM, Schuep W, Klag MJ. Serum antioxidants and myocardial infarction. Are low levels of carotenoids and alpha-tocopherol risk factors for myocardial infarction? Circulation. 1994;90(3):1154-1161. (PubMed)
132. Ito Y, Kurata M, Suzuki K, Hamajima N, Hishida H, Aoki K. Cardiovascular disease mortality and serum carotenoid levels: a Japanese population-based follow-up study. J Epidemiol. 2006;16(4):154-160. (PubMed)
133. Buijsse B, Feskens EJ, Kwape L, Kok FJ, Kromhout D. Both alpha- and beta-carotene, but not tocopherols and vitamin C, are inversely related to 15-year cardiovascular mortality in Dutch elderly men. J Nutr. 2008;138(2):344-350. (PubMed)
134. Sesso HD, Buring JE, Norkus EP, Gaziano JM. Plasma lycopene, other carotenoids, and retinol and the risk of cardiovascular disease in men. Am J Clin Nutr. 2005;81(5):990-997. (PubMed)
135. Hak AE, Stampfer MJ, Campos H, et al. Plasma carotenoids and tocopherols and risk of myocardial infarction in a low-risk population of US male physicians. Circulation. 2003;108(7):802-807. (PubMed)
136. Evans RW, Shaten BJ, Day BW, Kuller LH. Prospective association between lipid soluble antioxidants and coronary heart disease in men. The Multiple Risk Factor Intervention Trial. Am J Epidemiol. 1998;147(2):180-186. (PubMed)
137. Sahyoun NR, Jacques PF, Russell RM. Carotenoids, vitamins C and E, and mortality in an elderly population. Am J Epidemiol. 1996;144(5):501-511. (PubMed)
138. Wang Y, Chung SJ, McCullough ML, et al. Dietary carotenoids are associated with cardiovascular disease risk biomarkers mediated by serum carotenoid concentrations. J Nutr. 2014;144(7):1067-1074. (PubMed)
139. Li Z, Chen J, Zhang D. Association between dietary carotenoid intakes and hypertension in adults: National Health and Nutrition Examination Survey 2007-2014. J Hypertens. 2019;37(12):2371-2379. (PubMed)
140. Leermakers ET, Darweesh SK, Baena CP, et al. The effects of lutein on cardiometabolic health across the life course: a systematic review and meta-analysis. Am J Clin Nutr. 2016;103(2):481-494. (PubMed)
141. Tierney AC, Rumble CE, Billings LM, George ES. Effect of dietary and supplemental lycopene on cardiovascular risk factors: a systematic review and meta-analysis. Adv Nutr. 2020;11(6):1453-1488. (PubMed)
142. Rimm EB, Stampfer MJ, Ascherio A, Giovannucci E, Colditz GA, Willett WC. Vitamin E consumption and the risk of coronary heart disease in men. N Engl J Med. 1993;328(20):1450-1456. (PubMed)
143. Gaziano JM, Manson JE, Branch LG, Colditz GA, Willett WC, Buring JE. A prospective study of consumption of carotenoids in fruits and vegetables and decreased cardiovascular mortality in the elderly. Ann Epidemiol. 1995;5(4):255-260. (PubMed)
144. Osganian SK, Stampfer MJ, Rimm E, Spiegelman D, Manson JE, Willett WC. Dietary carotenoids and risk of coronary artery disease in women. Am J Clin Nutr. 2003;77(6):1390-1399. (PubMed)
145. Greenberg ER, Baron JA, Karagas MR, et al. Mortality associated with low plasma concentration of beta carotene and the effect of oral supplementation. JAMA. 1996;275(9):699-703. (PubMed)
146. Omenn GS, Goodman GE, Thornquist MD, et al. Effects of a combination of beta carotene and vitamin A on lung cancer and cardiovascular disease. N Engl J Med. 1996;334(18):1150-1155. (PubMed)
147. Yang J, Zhang Y, Na X, Zhao A. beta-Carotene supplementation and risk of cardiovascular disease: a systematic review and meta-analysis of randomized controlled trials. Nutrients. 2022;14(6):1284. (PubMed)
148. Voutilainen S, Nurmi T, Mursu J, Rissanen TH. Carotenoids and cardiovascular health. Am J Clin Nutr. 2006;83(6):1265-1271. (PubMed)
149. Gajendragadkar PR, Hubsch A, Maki-Petaja KM, Serg M, Wilkinson IB, Cheriyan J. Effects of oral lycopene supplementation on vascular function in patients with cardiovascular disease and healthy volunteers: a randomised controlled trial. PLoS One. 2014;9(6):e99070. (PubMed)
150. Stangl V, Kuhn C, Hentschel S, et al. Lack of effects of tomato products on endothelial function in human subjects: results of a randomised, placebo-controlled cross-over study. Br J Nutr. 2011;105(2):263-267. (PubMed)
151. Kim JY, Paik JK, Kim OY, et al. Effects of lycopene supplementation on oxidative stress and markers of endothelial function in healthy men. Atherosclerosis. 2011;215(1):189-195. (PubMed)
152. Xaplanteris P, Vlachopoulos C, Pietri P, et al. Tomato paste supplementation improves endothelial dynamics and reduces plasma total oxidative status in healthy subjects. Nutr Res. 2012;32(5):390-394. (PubMed)
153. Ozaki K, Okamoto M, Fukasawa K, et al. Daily intake of beta-cryptoxanthin prevents bone loss by preferential disturbance of osteoclastic activation in ovariectomized mice. J Pharmacol Sci. 2015;129(1):72-77. (PubMed)
154. Sahni S, Hannan MT, Blumberg J, Cupples LA, Kiel DP, Tucker KL. Inverse association of carotenoid intakes with 4-y change in bone mineral density in elderly men and women: the Framingham Osteoporosis Study. Am J Clin Nutr. 2009;89(1):416-424. (PubMed)
155. Sahni S, Hannan MT, Blumberg J, Cupples LA, Kiel DP, Tucker KL. Protective effect of total carotenoid and lycopene intake on the risk of hip fracture: a 17-year follow-up from the Framingham Osteoporosis Study. J Bone Miner Res. 2009;24(6):1086-1094. (PubMed)
156. Dai Z, Wang R, Ang LW, Low YL, Yuan JM, Koh WP. Protective effects of dietary carotenoids on risk of hip fracture in men: the Singapore Chinese Health Study. J Bone Miner Res. 2014;29(2):408-417. (PubMed)
157. Sugiura M, Nakamura M, Ogawa K, Ikoma Y, Yano M. High serum carotenoids associated with lower risk for bone loss and osteoporosis in post-menopausal Japanese female subjects: prospective cohort study. PLoS One. 2012;7(12):e52643. (PubMed)
158. Hayhoe RPG, Lentjes MAH, Mulligan AA, Luben RN, Khaw KT, Welch AA. Carotenoid dietary intakes and plasma concentrations are associated with heel bone ultrasound attenuation and osteoporotic fracture risk in the European Prospective Investigation into Cancer and Nutrition (EPIC)-Norfolk cohort. Br J Nutr. 2017;117(10):1439-1453. (PubMed)
159. Kang JH, Ascherio A, Grodstein F. Fruit and vegetable consumption and cognitive decline in aging women. Ann Neurol. 2005;57(5):713-720. (PubMed)
160. Morris MC, Evans DA, Tangney CC, Bienias JL, Wilson RS. Associations of vegetable and fruit consumption with age-related cognitive change. Neurology. 2006;67(8):1370-1376. (PubMed)
161. Christensen K, Gleason CE, Mares JA. Dietary carotenoids and cognitive function among US adults, NHANES 2011-2014. Nutr Neurosci. 2020;23(7):554-562. (PubMed)
162. Edwards CG, Walk AM, Thompson SV, et al. Dietary lutein plus zeaxanthin and choline intake is interactively associated with cognitive flexibility in middle-adulthood in adults with overweight and obesity. Nutr Neurosci. 2022;25(7):1437-1452. (PubMed)
163. Feeney J, O'Leary N, Moran R, et al. Plasma lutein and zeaxanthin are associated with better cognitive function across multiple domains in a large population-based sample of older adults: findings from the Irish Longitudinal Study on Aging. J Gerontol A Biol Sci Med Sci. 2017;72(10):1431-1436. (PubMed)
164. Johnson EJ, Vishwanathan R, Johnson MA, et al. Relationship between serum and brain carotenoids, alpha-tocopherol, and retinol concentrations and cognitive performance in the oldest old from the Georgia Centenarian Study. J Aging Res. 2013;2013:951786. (PubMed)
165. Vishwanathan R, Kuchan MJ, Sen S, Johnson EJ. Lutein and preterm infants with decreased concentrations of brain carotenoids. J Pediatr Gastroenterol Nutr. 2014;59(5):659-665. (PubMed)
166. Feeney J, Finucane C, Savva GM, et al. Low macular pigment optical density is associated with lower cognitive performance in a large, population-based sample of older adults. Neurobiol Aging. 2013;34(11):2449-2456. (PubMed)
167. Renzi LM, Dengler MJ, Puente A, Miller LS, Hammond BR, Jr. Relationships between macular pigment optical density and cognitive function in unimpaired and mildly cognitively impaired older adults. Neurobiol Aging. 2014;35(7):1695-1699. (PubMed)
168. Kelly D, Coen RF, Akuffo KO, et al. Cognitive function and its relationship with macular pigment optical density and serum concentrations of its constituent carotenoids. J Alzheimers Dis. 2015;48(1):261-277. (PubMed)
169. Johnson EJ, McDonald K, Caldarella SM, Chung HY, Troen AM, Snodderly DM. Cognitive findings of an exploratory trial of docosahexaenoic acid and lutein supplementation in older women. Nutr Neurosci. 2008;11(2):75-83. (PubMed)
170. Renzi-Hammond LM, Bovier ER, Fletcher LM, et al. Effects of a lutein and zeaxanthin intervention on cognitive function: a randomized, double-masked, placebo-controlled trial of younger healthy adults. Nutrients. 2017;9(11):1246. (PubMed)
171. Power R, Coen RF, Beatty S, et al. Supplemental retinal carotenoids enhance memory in healthy individuals with low levels of macular pigment in a randomized, double-blind, placebo-controlled clinical trial. J Alzheimers Dis. 2018;61(3):947-961. (PubMed)
172. Chew EY, Clemons TE, Agron E, et al. Effect of omega-3 fatty acids, lutein/zeaxanthin, or other nutrient supplementation on cognitive function: the AREDS2 randomized clinical trial. JAMA. 2015;314(8):791-801. (PubMed)
173. Yeum KJ, Russell RM. Carotenoid bioavailability and bioconversion. Annu Rev Nutr. 2002;22:483-504. (PubMed)
174. Gartner C, Stahl W, Sies H. Lycopene is more bioavailable from tomato paste than from fresh tomatoes. Am J Clin Nutr. 1997;66(1):116-122. (PubMed)
175. Stahl W, Sies H. Uptake of lycopene and its geometrical isomers is greater from heat-processed than from unprocessed tomato juice in humans. J Nutr. 1992;122(11):2161-2166. (PubMed)
176. US Department of Agriculture, Agricultural Research Service. FoodData Central. Available at https://fdc.nal.usda.gov/. Accessed 9/28/2023.
177. Clinton SK. Lycopene: chemistry, biology, and implications for human health and disease. Nutr Rev. 1998;56(2 Pt 1):35-51. (PubMed)
178. Hendler SS, Rorvik DM, eds. PDR for Nutritional Supplements. 2nd ed. Thomson Reuters; 2008.
179. Tudor C, Pintea A. A brief overview of dietary zeaxanthin occurrence and bioaccessibility. Molecules. 2020;25(18):4067. (PubMed)
180. Tanvetyanon T, Bepler G. Beta-carotene in multivitamins and the possible risk of lung cancer among smokers versus former smokers: a meta-analysis and evaluation of national brands. Cancer. 2008;113(1):150-157. (PubMed)
181. Bowen PE, Herbst-Espinosa SM, Hussain EA, Stacewicz-Sapuntzakis M. Esterification does not impair lutein bioavailability in humans. J Nutr. 2002;132(12):3668-3673. (PubMed)
182. Solomons NW. Vitamin A and carotenoids. In: Bowman BA, Russell RM, eds. Present Knowledge in Nutrition. 8th ed. Washington, D.C.: ILSI Press; 2001:127-145.
183. Shao A, Hathcock JN. Risk assessment for the carotenoids lutein and lycopene. Regul Toxicol Pharmacol. 2006;45(3):289-298. (PubMed)
184. Choi RY, Chortkoff SC, Gorusupudi A, Bernstein PS. Crystalline maculopathy associated with high-dose lutein supplementation. JAMA Ophthalmol. 2016;134(12):1445-1448. (PubMed)
185. Tan KML, Chee J, Lim KLM, et al. Safety, tolerability, and pharmacokinetics of beta-cryptoxanthin supplementation in healthy women: a double-blind, randomized, placebo-controlled clinical trial. Nutrients. 2023;15(10):2325. (PubMed)
186. Ehrenfeld M, Levy M, Sharon P, Rachmilewitz D, Eliakim M. Gastrointestinal effects of long-term colchicine therapy in patients with recurrent polyserositis (familial mediterranean fever). Dig Dis Sci. 1982;27(8):723-727. (PubMed)
187. Natural-Medicines. Beta carotene/Drug interactions - Professional handout; 2016.
188. Brown BG, Zhao XQ, Chait A, et al. Simvastatin and niacin, antioxidant vitamins, or the combination for the prevention of coronary disease. N Engl J Med. 2001;345(22):1583-1592. (PubMed)
189. Collins R, Peto R, Armitage J. The MRC/BHF Heart Protection Study: preliminary results. Int J Clin Pract. 2002;56(1):53-56. (PubMed)
190. Koonsvitsky BP, Berry DA, Jones MB, et al. Olestra affects serum concentrations of alpha-tocopherol and carotenoids but not vitamin D or vitamin K status in free-living subjects. J Nutr. 1997;127(8 Suppl):1636S-1645S. (PubMed)
191. Thornquist MD, Kristal AR, Patterson RE, et al. Olestra consumption does not predict serum concentrations of carotenoids and fat-soluble vitamins in free-living humans: early results from the sentinel site of the olestra post-marketing surveillance study. J Nutr. 2000;130(7):1711-1718. (PubMed)
192. Neuhouser ML, Rock CL, Kristal AR, et al. Olestra is associated with slight reductions in serum carotenoids but does not markedly influence serum fat-soluble vitamin concentrations. Am J Clin Nutr. 2006;83(3):624-631. (PubMed)
193. Katan MB, Grundy SM, Jones P, Law M, Miettinen T, Paoletti R. Efficacy and safety of plant stanols and sterols in the management of blood cholesterol levels. Mayo Clin Proc. 2003;78(8):965-978. (PubMed)
194. Weststrate JA, Meijer GW. Plant sterol-enriched margarines and reduction of plasma total- and LDL-cholesterol concentrations in normocholesterolaemic and mildly hypercholesterolaemic subjects. Eur J Clin Nutr. 1998;52(5):334-343. (PubMed)
195. Noakes M, Clifton P, Ntanios F, Shrapnel W, Record I, McInerney J. An increase in dietary carotenoids when consuming plant sterols or stanols is effective in maintaining plasma carotenoid concentrations. Am J Clin Nutr. 2002;75(1):79-86. (PubMed)
196. Leo MA, Lieber CS. Alcohol, vitamin A, and beta-carotene: adverse interactions, including hepatotoxicity and carcinogenicity. Am J Clin Nutr. 1999;69(6):1071-1085. (PubMed)
197. Albanes D, Heinonen OP, Taylor PR, et al. Alpha-tocopherol and beta-carotene supplements and lung cancer incidence in the alpha-tocopherol, beta-carotene cancer prevention study: effects of base-line characteristics and study compliance. J Natl Cancer Inst. 1996;88(21):1560-1570. (PubMed)
198. van den Berg H. Carotenoid interactions. Nutr Rev. 1999;57(1):1-10. (PubMed)
199. Micozzi MS, Brown ED, Edwards BK, et al. Plasma carotenoid response to chronic intake of selected foods and beta-carotene supplements in men. Am J Clin Nutr. 1992;55(6):1120-1125. (PubMed)
200. Kostic D, White WS, Olson JA. Intestinal absorption, serum clearance, and interactions between lutein and beta-carotene when administered to human adults in separate or combined oral doses. Am J Clin Nutr. 1995;62(3):604-610. (PubMed)
201. Albanes D, Virtamo J, Taylor PR, Rautalahti M, Pietinen P, Heinonen OP. Effects of supplemental beta-carotene, cigarette smoking, and alcohol consumption on serum carotenoids in the Alpha-Tocopherol, Beta-Carotene Cancer Prevention Study. Am J Clin Nutr. 1997;66(2):366-372. (PubMed)
202. Nierenberg DW, Dain BJ, Mott LA, Baron JA, Greenberg ER. Effects of 4 y of oral supplementation with beta-carotene on serum concentrations of retinol, tocopherol, and five carotenoids. Am J Clin Nutr. 1997;66(2):315-319. (PubMed)
203. Wahlqvist ML, Wattanapenpaiboon N, Macrae FA, Lambert JR, MacLennan R, Hsu-Hage BH. Changes in serum carotenoids in subjects with colorectal adenomas after 24 mo of beta-carotene supplementation. Australian Polyp Prevention Project Investigators. Am J Clin Nutr. 1994;60(6):936-943. (PubMed)





