Contents
Although choline is not by strict definition a vitamin, it is an essential nutrient. Despite the fact that humans can synthesize it in small amounts, choline must be consumed in the diet to maintain health. The majority of the body's choline is found in specialized fat molecules known as phospholipids, the most common of which is called phosphatidylcholine (1).
Choline and compounds derived from choline (i.e., metabolites) serve a number of vital biological functions (Figure 1) (1).

Choline is used in the synthesis of certain phospholipids (phosphatidylcholine and sphingomyelin) that are essential structural components of cell membranes. Phosphatidylcholine accounts for about 95% of total choline in tissues (2). This phospholipid can be synthesized from dietary choline via the cytidine diphosphocholine (CDP-choline) pathway or through the methylation of another phospholipid, phosphatidylethanolamine (Figure 2) (3). Sphingomyelin is a type of sphingosine-containing phospholipid (sphingolipid) that is synthesized by the transfer of a phosphocholine residue from a phosphatidylcholine to a ceramide (Figure 3). Sphingomyelin is found in cell membranes and in the fatty sheath that envelops myelinated nerve fibers.



The choline-containing phospholipids, phosphatidylcholine, and sphingomyelin, are precursors for the intracellular messenger molecules, diacylglycerol and ceramide. Specifically, sphingomyelinases (also known as sphingomyelin phosphodiesterases) catalyze the cleavage of sphingomyelin, generating phosphocholine and ceramide. Diacylglycerol is released by the degradation of phosphatidylcholine by phospholipases. Other choline metabolites known to be cell-signaling molecules include platelet-activating factor (PAF) and sphingophosphocholine.
Choline is a precursor for acetylcholine, an important neurotransmitter synthesized by cholinergic neurons and involved in muscle control, circadian rhythm, memory, and many other neuronal functions. Choline acetyltransferase catalyzes the acetylation of choline to acetylcholine, and acetylcholine esterase hydrolyzes acetylcholine to choline and acetate (4). CDP-choline (citicoline) administration was also found to stimulate the synthesis and release of a family of neurotransmitters derived from tyrosine (i.e., the catecholamines, including noradrenaline, adrenaline, and dopamine) (5). Of note, non-neuronal cells of various tissues and organ systems also synthesize and release acetylcholine, which then binds and stimulates cholinergic receptors on target cells (reviewed in 6).
Fat and cholesterol consumed in the diet are transported to the liver by lipoproteins called chylomicrons. In the liver, fat and cholesterol are packaged into lipoproteins called very-low-density lipoproteins (VLDL) for transport in the bloodstream to extrahepatic tissues. Phosphatidylcholine synthesis by the phosphatidylethanolamine N-methyltransferase (PEMT) pathway is required for VLDL assembly and secretion from the liver (7, 8). Polymorphisms of the PEMT gene increase the dietary requirements of choline (9). Without adequate phosphatidylcholine, fat and cholesterol accumulate in the liver (see Deficiency).
Choline may be oxidized in the liver and kidney to form a metabolite called betaine via a two-step enzymatic reaction. In the mitochondrial inner membrane, flavin adenine dinucleotide (FAD)-dependent choline oxidase catalyzes the conversion of choline to betaine aldehyde, which is then converted to betaine by betaine aldehyde dehydrogenase in either the mitochondrial matrix or the cytosol (2). Betaine is a source of up to 60% of the methyl (CH3) groups required for the methylation of homocysteine (10). Betaine homocysteine methyltransferase (BHMT) uses betaine as a methyl donor to convert homocysteine to methionine in one-carbon metabolism (Figure 4). The ubiquitous vitamin B12-dependent methionine synthase (MS) enzyme also catalyzes the re-methylation of homocysteine, using the folate derivative, 5-methyltetrahydrofolate, as a methyl donor (see Nutrient interactions). Elevated concentrations of homocysteine in the blood have been associated with increased risk of cardiovascular disease (11).

The conversion of choline to betaine is irreversible. Betaine is an osmolyte that regulates cell volume and protect cell integrity against osmotic stress (especially in the kidney). Osmotic stress has been associated with a reduced BHMT expression such that the role of betaine in osmoregulation may be temporarily prioritized over its function as a methyl donor (2).
Men and women fed intravenously (IV) with solutions that contained adequate methionine and folate but lacked choline have been found to develop a condition called nonalcoholic fatty liver disease (NAFLD) and signs of liver damage that resolved when choline was provided (12). The occurrence of NAFLD is usually associated with the co-presentation of metabolic disorders, including obesity, dyslipidemia, insulin resistance, and hypertension, in subjects with metabolic syndrome. NAFLD is estimated to progress to a more severe condition called nonalcoholic steatohepatitis (NASH) in about one-third of NAFLD patients, as well as to increase the risk of cirrhosis and liver cancer (13).
Because phosphatidylcholine is required in the synthesis of very-low-density lipoprotein (VLDL) particles (see Function), choline deficiency results in impaired VLDL secretion and accumulation of fat in the liver (steatosis), ultimately leading to liver damage. Because low-density lipoprotein (LDL) particles are formed from VLDL particles, choline-deficient individuals also show reduced blood concentrations of LDL-cholesterol (14). Abnormally elevated biomarkers of organ dysfunction in the blood, including creatine phosphokinase, aspartate aminotransferase, and alanine aminotransferase, are corrected upon choline repletion. Choline deficiency-induced organ dysfunction has also been associated with increased DNA damage and apoptosis in circulating lymphocytes (15). In the liver, the accumulation of lipids is thought to impair mitochondrial function, thus reducing fatty acid oxidation and increasing the production of reactive oxygen species (ROS) that trigger lipid peroxidation, DNA damage, and apoptosis. Further, oxidative stress is thought to be responsible for prompting inflammatory processes that can lead to the progression of NAFLD to NASH and cirrhosis (end-stage liver disease) (16).
An intervention study in 57 healthy adults who were fed choline-deficient diets under controlled conditions found that 77% of men, 80% of postmenopausal women, and 44% of premenopausal women developed fatty liver, liver damage, and/or muscle damage (17). These signs of organ dysfunction resolved upon choline reintroduction in the diet. Because estrogen stimulates the endogenous synthesis of phosphatidylcholine via the phosphatidylethanolamine N-methyltransferase (PEMT) pathway, premenopausal women may be less likely to develop signs of choline deficiency in response to a low-choline diet compared to postmenopausal women (18, 19). Further, a notable single nucleotide polymorphism (SNP; rs12325817) of the PEMT gene, which may affect the expression and/or activity of the PEMT enzyme, is thought to increase the susceptibility to choline deficiency-induced organ dysfunction (18). Additional genetic polymorphisms occurring in choline and one-carbon metabolic pathways may alter the dietary requirement for choline and thus increase the likelihood of developing signs of deficiency when choline intake is inadequate (9, 20, 21).
Of note, the composition of one’s intestinal microbiota has been identified as a potential predictor of susceptibility to choline deficiency-induced NAFLD (22). Intestinal microbiota-dependent metabolism of dietary phosphatidylcholine might also be involved in the pathogenesis of cardiovascular disease (see Safety) (23, 24).
For more information on fatty liver diseases, see the section on Disease Prevention.
Daily intake recommendations are higher during pregnancy and lactation (see The AI), placing pregnant and lactating individuals at increased risk of choline inadequacy (25, 26). Vegetarians may also be at increased risk of choline inadequacy (26).
Patients with cystic fibrosis who have pancreatic insufficiency are at risk of choline depletion due to increased fecal losses (27). Circulating choline has been directly correlated with lung function in cystic fibrosis patients (28), and results of a pilot study in 10 patients suggested that choline supplementation may have utility in improving clinical outcomes in cystic fibrosis patients (29).
Together with several B-vitamins (i.e., folate, vitamin B12, vitamin B6, and riboflavin), choline is required for the metabolism of nucleic acids and amino acids, and for the generation of the universal methyl group donor, S-adenosylmethionine (SAM) (Figure 4). SAM is synthesized from the essential amino acid, methionine. Three molecules of SAM are required for the methylation reaction that converts phosphatidylethanolamine into phosphatidylcholine (Figure 2). Once SAM donates a methyl group it becomes S-adenosylhomocysteine (SAH), which is then metabolized to homocysteine. Homocysteine can be converted back to methionine in a reaction catalyzed by vitamin B12-dependent methionine synthase, which requires 5-methyltetrahydrofolate (5-meTHF) as a methyl donor. Alternately, betaine (a metabolite of choline) is used as the methyl donor for the methylation of homocysteine to methionine by the enzyme, betaine-homocysteine methyltransferase (BHMT) (1). Homocysteine can also be metabolized to cysteine via the vitamin B6-dependent transsulfuration pathway (Figure 4).
Thus, the human requirement for choline is especially influenced by the relationship between choline and other methyl group donors, such as folate and S-adenosylmethionine. A low intake of folate leads to an increased demand for choline-derived metabolite, betaine. Moreover, the de novo synthesis of phosphatidylcholine is not sufficient to maintain adequate choline nutritional status when dietary intakes of folate and choline are low (30). Conversely, the demand for folate is increased when dietary supply for choline is limited (31).
In 1998, the Food and Nutrition Board (FNB) of the Institute of Medicine (now the National Academy of Medicine) established a dietary reference intake (DRI) for choline (32). The FNB felt the existing scientific evidence was insufficient to calculate an RDA for choline, so they set an Adequate Intake (AI; Table 1). The main criterion for establishing the AI for choline was the prevention of liver damage. Yet, common polymorphisms in genes involved in choline or folate metabolism alter one’s susceptibility to choline deficiency and thus may affect dietary requirements for choline (see Deficiency) (18, 20, 33, 34).
A large body of research indicates that even moderately elevated levels of homocysteine in the blood increase the risk of cardiovascular disease (11). The most common cause of a myocardial infarction or a stroke is the rupture of atherosclerotic plaques in arterial walls causing blood clot formation (thrombogenesis). High homocysteine concentrations may promote the development of atherosclerosis (atherogenesis) and thrombogenesis via mechanisms involving oxidative stress and endothelial dysfunction, inflammation, abnormal blood coagulation, and disordered lipid metabolism (reviewed in 35).
Once formed from dietary methionine, homocysteine can be catabolized to cysteine via the transsulfuration pathway or re-methylated to methionine (Figure 4). Folate and choline are involved in alternate pathways that catalyze the re-methylation of homocysteine (see Nutrient interactions). Specifically, choline is the precursor of betaine, which provides a methyl group for the conversion of homocysteine to methionine via the enzyme, betaine-homocysteine methyltransferase (BHMT). While the amount of homocysteine in the blood is regulated by several nutrients, including folate and choline, conditions that cause damage to the liver like nonalcoholic steatohepatitis (NASH) may also affect homocysteine metabolism (36).
Because both folate- and choline-dependent metabolic pathways catalyze the re-methylation of homocysteine, dietary intakes of both nutrients need to be considered when the association between homocysteine concentrations and cardiovascular disease is assessed. Yet, despite its relevance, the relationship of betaine and choline to homocysteine metabolism has been only lightly investigated in humans, essentially because the choline content of foods could not be accurately measured until fairly recently. In preliminary intervention studies, pharmacologic doses of betaine (1,500 to 6,000 mg/day) were found to reduce blood homocysteine concentrations in a small number of volunteers with normal-to-mildly elevated homocysteine concentrations (37-40). Yet, in a cross-sectional analysis of a large cohort of 16,165 women (ages, 49-79 years), lower betaine doses in the range of dietary intakes were not found to be correlated with homocysteine concentrations (41). This study also showed that levels of choline intake were inversely associated with homocysteine concentrations in the blood. However, an eight-year follow-up study of the cohort failed to show any difference in cardiovascular risk between women in the upper versus bottom quartile of dietary choline intakes (>329 mg/day vs. ≤266 mg/day) (41).
More recent prospective cohort studies on the association of dietary choline or betaine and cardiovascular disease have been mixed. In the Jackson Heart Study, which followed 3,924 African Americans for nine years, higher dietary choline intakes were associated with a reduced risk of stroke (42). In the Nurses’ Health Study (80,978 women) and the Health Professionals Follow-up Study (39,434 men), higher dietary intakes of phosphatidylcholine were linked to a higher risk of cardiovascular disease-related mortality (43). Higher total daily choline intakes were linked to a higher risk of cardiometabolic mortality (i.e., death attributed to cardiovascular disease or type 2 diabetes mellitus) in large prospective cohorts of three ethnic groups: blacks (n=49,858), whites (n=23,766), and Chinese (n=134,001) (44). In this study, higher betaine intakes were associated with a higher risk of cardiometabolic death in the Chinese cohort only (44). However, no associations between choline or betaine intakes and cardiovascular-related mortality were found in a cohort study of Japanese adults (n=29,079) (45). Several other prospective cohort studies have failed to find an association between choline intake and cardiovascular disease endpoints (41, 46-48).
Convincing evidence that increased dietary intake of choline or betaine could benefit cardiovascular health through lowering homocysteine concentrations in the blood is presently lacking.
A 1995 study had found that elevated blood homocysteine concentrations in patients who experienced a vascular occlusion were associated with higher urinary excretion of betaine, rather than with reduced intake of choline or betaine or diminished activity of BHMT (49). In a prospective study, high urinary betaine excretion was also associated with increased risk of heart failure in 325 subjects without diabetes mellitus who had been hospitalized for acute coronary syndrome (50). In the same study, both top and bottom quintiles of plasma betaine concentrations were associated with an increased risk of secondary acute myocardial infarction. The findings of another prospective study (the Hordaland Health Study) that followed 7,045 healthy adults (ages, 47-49 years and 71-74 years) suggested that high choline and low betaine plasma concentrations were associated with an unfavorable cardiovascular risk profile (51). Indeed, plasma choline was positively associated with a number of cardiovascular risk factors, such as BMI, percentage body fat, waist circumference, and serum triglycerides, and inversely associated with HDL-cholesterol. On the contrary, plasma betaine was positively correlated to HDL-cholesterol and inversely associated with the above-mentioned risk factors, as well as with systolic and diastolic blood pressure.
More recent studies suggest that the blood concentration of trimethylamine N-oxide (TMAO), generated from trimethylamine-containing nutrients like dietary choline, rather than that of choline, might influence the risk of cardiovascular events (see Safety). However, the association between high blood levels of TMAO and atherosclerosis was not observed in the Coronary Artery Risk Development in Young Adults Study (CARDIA) (52)
It is not yet clear whether concentrations of choline, betaine, and/or TMAO in the blood can predict the risk for cardiovascular disease.
While a choline-deficient diet results in organ dysfunction and nonalcoholic fatty liver disease (NAFLD) (see Deficiency; 17), it is not known whether suboptimal dietary choline intakes in healthy subjects may contribute to an increased risk for NAFLD. A cross-sectional analysis of two large prospective studies conducted in China — the Shanghai Women’s Health Study and the Shanghai Men’s Health Study — including 56,195 people (ages, 40-75 years), was conducted to assess the association between dietary choline intakes and self-reported diagnosis of fatty liver disease (53). The highest versus lowest quintile of choline intake (412 mg/day vs. 179 mg/day) was associated with a 28% lower risk of fatty liver disease in normal-weight women, but no association was found in overweight or obese women or in men. Another cross-sectional study of 664 individuals with NAFLD or nonalcoholic steatohepatitis (NASH) also reported that disease severity was inversely correlated with dietary choline intakes in postmenopausal women, but not in premenopausal women, men, or children (54). Moreover, in a US national cross-sectional survey of 20,643 adults, higher dietary intakes of choline were associated with a more favorable profile of liver enzymes and with a lower risk of developing NAFLD (NHANES 2005-2012) (55).
In animal models, dietary choline deficiency has been associated with an increased incidence of spontaneous liver cancer (hepatocellular carcinoma) and increased sensitivity to carcinogenic chemicals (10). A number of mechanisms have been proposed to contribute to the cancer-promoting effects of choline deficiency: (1) enhanced liver cell regeneration and tissue sensitivity to chemical insults; (2) altered expression of numerous genes regulating cell proliferation, differentiation, DNA repair, and apoptosis due to improper DNA methylation; (3) increased likelihood of DNA damage caused by mitochondrial dysfunction-induced oxidative stress; and (4) activated protein kinase C-mediated cell-signaling cascade, eventually leading to an increase in liver cell apoptosis (2). Yet, it is not known whether choline deficiency can increase the susceptibility to cancer in humans (2).
It is known that folate is critical for normal embryonic development, and maternal supplementation with folic acid decreases the incidence of neural tube defects (NTDs) (56). NTDs include various malformations, such as lesions of the brain (e.g., anencephaly, encephalocele) or lesions of the spine (spina bifida), which are devastating and usually incompatible with life (57). These defects occur between the 21st and 28th days after conception, a time when many women do not realize that they are pregnant (58). While the protective effect of folate against NTD is well established, only a few studies have investigated the role of other methyl group donors, including choline and betaine, in the occurrence of NTDs. A case-control study (424 NTD cases and 440 controls) found that women in the highest versus lowest quartile of periconceptual choline intake (>498.46 mg/day vs. ≤290.41 mg/day) had a 51% lower risk of an NTD-affected pregnancy (59). However, more recent studies have failed to find an inverse relationship between maternal choline intake and risk of NTDs (60-63).
A case-control study (80 NTD-affected pregnancy and 409 controls) in US women found that the lowest concentrations of serum choline (<2.49 mmol/L) during mid-pregnancy were associated with a 2.4-fold higher risk of NTDs (64). Finally, a more recent study, including 71 NTD-affected pregnancies, 214 pregnancies with non NTD malformations, 98 normal pregnancies in women with prior NTD-affected pregnancies, and 386 normal pregnancies, found no associations between maternal blood concentrations of choline during pregnancy, choline- and folate-related genetic variants, and risk of NTDs (65). However, it is important to note that circulating choline concentrations do not accurately reflect dietary intake of choline.
In a recent meta-analysis that pooled the results of five case-control studies (59, 60, 62-64), including 1,131 NTD-affected pregnancies and 4,439 healthy controls, lower dietary choline intakes or lower serum concentrations of choline were associated with a 36% higher risk of NTDs compared to higher levels (95% CI, 1.11, 1.67) (66).
Randomized controlled trials of choline supplementation throughout the periconceptional period would been needed to determine whether choline has a protective effect against NTDs.
Increased dietary intake of cytidine 5’-diphosphocholine (CDP-choline or citicoline, a precursor of phosphatidylcholine; Figure 2) very early in life can diminish the severity of memory deficits in aged rats (67). Choline supplementation of the mothers of unborn rats, as well as rat pups during the first month of life, led to improved performance in spatial memory tests months after choline supplementation had been discontinued (68). A review by McCann et al. discusses the experimental evidence from rodent studies regarding the availability of choline during prenatal development and cognitive function in the offspring (69).
Because of the importance of DNA methylation in normal brain development, neuronal functions, and cognitive processes (70), methyl donor nutrients like choline are essential for optimal brain functioning. However, clinical evidence to determine whether findings in rodent studies are applicable to humans is currently limited. The analysis of the Seychelles Child Development Nutrition Cohort study reported a lack of an association between plasma concentrations of choline and its related metabolites and cognitive abilities in 256 five-year-old children. Only plasma betaine concentrations were found to be positively correlated with preschool language test scores (71). Yet, because circulating concentrations of choline are not directly related to dietary choline intakes, the study could not evaluate whether maternal choline intakes influence children’s brain development.
Project Viva is an ongoing prospective study that has examined the relationship between daily intakes of methyl donor nutrients in 1,210 women during pregnancy and child cognition at three and seven years postpartum. Maternal intake of choline during the first and/or second trimester of pregnancy was not correlated with measures of cognitive performance in children at age 3 years (72). Another report of the study indicated that upper versus lower quartile of maternal choline intakes during the second trimester of pregnancy (median intakes, 392 mg/day vs. 260 mg/day) was significantly associated with higher visual memory scores in children ages 7 years old (73). In addition, a small randomized, double-blind, placebo-controlled trial in 99 pregnant women (ages, 21-41 years old) evaluated the effect of choline supplementation during pregnancy and lactation on infants’ cognitive function at ages 10 and 12 months (74). The results indicated that maternal choline supplementation (750 mg/day of choline in the form of phosphatidylcholine) from 18 weeks of gestation to 3 months’ postpartum provided no cognitive benefits in children regarding short-term visuospatial memory, long-term episodic memory, and language and global development (74). In a randomized, double-blind, controlled-feeding study in 24 pregnant women, maternal choline intake of 930 mg/day (~2x the AI) throughout the third trimester of pregnancy improved measures of information-processing speed (i.e., reaction times) and visuospatial memory in infants compared to choline intakes of 480 mg/day (75). Choline intake in this study was from both dietary and supplemental sources: 380 mg/day of choline from dietary sources and either 100 mg/day or 550 mg/day from supplemental choline chloride (75). Follow-up of the offspring (N=20) at 7 years of age indicated that children of mothers who consumed 930 mg/day of choline had higher measures of sustained attention compared to those of mothers who consumed 480 mg/day of choline (76), suggesting that prenatal choline supplementation during late pregnancy can confer cognitive benefits to the child.
Cognitive function, including the domains of memory, speed, and executive function, decline gradually with increasing age. The rate of cognitive decline is also influenced by modifiable risk factors like dietary habits. Deficiency in B-vitamins and elevated blood concentrations of homocysteine have been associated with cognitive impairments in the elderly. A recent meta-analysis of 14 randomized, placebo-controlled trials found that B-vitamin supplementation slowed cognitive decline in cognitive healthy older adults, as measured by score on the Mini-Mental State Examination, compared to placebo (77). However, less is known about dietary or supplemental choline specifically.
A few observational studies have examined choline intake and cognitive function in older adults, but these are cross-sectional in nature. The cross-sectional data analysis of a subgroup of 1,391 volunteers (ages, 36-83 years) from the large Framingham Heart Study Offspring cohort has indicated that dietary choline intake was positively associated with specific cognitive functions, namely verbal memory and visual memory (78). In a US national cross-sectional study of 2,393 older adults (≥60 years), total daily choline intakes (combined from diet and supplements) between 187.06 and 399.5 mg/day were associated with improved cognitive performance in three separate measures (assessing learning, processing speed, sustained attention, and working memory) compared to intakes less than 187.06 mg/day; however, the highest intakes of choline (>399.5 mg/day) were no different than the lowest intakes (<187.06 mg/day) in these measures (79).
Another cross-sectional study of 2,195 older individuals (ages, 70-74 years) from the Hordaland Health Study examined cognitive abilities and blood concentrations of various determinants of circulating homocysteine, including choline and betaine (80). Unlike betaine, high versus low plasma concentrations of free choline (>8.36 µM vs. ≤8.36 µM) were found to be significantly associated with a greater performance at cognitive tests assessing sensory motor speed, perceptual speed, executive function, and global cognition. However, in an earlier intervention study that enrolled 235 elderly individuals (mean age, 81 years old) with or without mild vitamin B12 deficiency, baseline concentrations of betaine — but not choline — were found to be positively correlated to test scores evaluating the cognitive domains of construction, sensory motor speed, and executive function (81).
More research is needed to determine the effect of choline on the developing brain and whether choline intakes above the RDA may be useful in the prevention of memory loss or dementia in older adults.
Neurodegenerative diseases, such as Alzheimer's disease (AD) and Parkinson’s disease (PD), are characterized by progressive cognitive decline and dementia. Dysfunctions in neurotransmitter signaling, affecting cholinergic and dopaminergic pathways in particular, have been involved in the occurrence of cognitive impairments. Deficits in acetylcholine and abnormal phospholipid metabolism have been reported in postmortem studies of the brains of AD patients (12). For these reasons, inhibitors of (acetyl) cholinesterase (which catalyzes the breakdown of acetylcholine) and large doses of lecithin (phosphatidylcholine) have been used to treat patients with dementia due to AD in hopes of raising the amount of acetylcholine available in the brain. While cholinesterase inhibitors have shown positive effects on cognitive functions and measures of clinical global state (82), a systematic review of randomized controlled trials did not find lecithin to be more beneficial than placebo in the treatment of patients with cognitive impairment, vascular dementia, AD, or mixed dementia (83). Limited data — mostly from case-control studies — are available to assess whether citicoline (CDP-choline) might improve cognitive performance in subjects with AD or PD, and studies to date have generally been labeled as poor in quality (84).
Optic neuropathies, including glaucoma, are associated with damage of the optic nerve and loss of visual function. In glaucoma, the progressive deterioration of the optic nerve is caused by loss of a specific neuronal population known as retinal ganglion cells (RGC), such that the condition has been classified as a neurodegenerative disease (85). Choline and its metabolites have a number of roles in supporting normal visual function, including retinal function (86).
In a small, double-blind, placebo-controlled study, the effect of citicoline was assessed in 24 subjects affected by open-angle glaucoma and treated with β-blockers. Patients were randomized to follow a therapeutic cycle for a total period of eight years: citicoline (1,000 mg/day, by intramuscular injection) or placebo (β-blockers alone) for a two-month period followed by a four-month washout period (87). Electrophysiological examinations were used to assess the extent of visual dysfunctions, including the simultaneous recordings of Pattern ElectroRetinoGrams (PERG) and Visual Evoked Potentials (VEP). Citicoline was found to enhance retinal function and neural conduction along post-retinal visual pathways, such that responses of the visual cortex to stimuli were significantly improved compared to placebo (87).
In a similar pilot trial, citicoline efficacy was assessed in 26 volunteers (mean age, 65.4 years) affected by another type of optic neuropathy known as non-arteritic anterior ischemic optic neuropathy (NAION). Oral citicoline (1,600 mg/day) was given for 60 days followed by 60 days of washout, and the therapeutic cycle was repeated once. Compared to placebo, citicoline was found to improve retinal function and post-retinal neural conduction, evidenced by PERG and VEP measures (88). Oral citicoline (four cycles of 500 mg/day for four months followed by a two-month washout period) was also found to significantly reduce the rate of visual field loss and the level of intraocular pressure in 41 patients with progressive glaucoma (89). Larger randomized controlled trials are needed to establish whether citicoline supplementation could be included in the medical treatment of glaucoma.
A few studies have also explored topical citicoline (i.e., eyedrops) as a potential treatment for glaucoma. One study in 56 patients with open-angle glaucoma found that such topically applied citicoline for four months improved retinal function and visual-related neural conduction when on top of β-blocker monotherapy to lower intraocular pressure (90). In a randomized, double-blind, placebo-controlled trial in 78 patients with progressive open-angle glaucoma (despite intraocular pressures of ≤18 mm Hg), use of citicoline eyedrops for three years blunted the decrease in retinal nerve fiber layer thickness compared to use of placebo eyedrops (91). This trial also found a trend (p=0.07) for a lower rate of disease progression with topical citicoline (91). While the results of these pilot studies are promising, large-scale randomized controlled trials are needed.
Cerebrovascular diseases (including stroke and sub-acute ischemic cerebrovascular disease) are the main cause of cognitive impairments in older people. Results from experimental studies have suggested that pharmacological doses of citicoline (CDP-choline) could enhance the metabolism of glucose and the biosynthesis of phospholipids and neurotransmitters, while limiting the degradation of phospholipids in neuronal membranes in models of ischemia and neurodegenerative diseases (reviewed in 92). Many short-term intervention studies in older individuals with vascular diseases have found that therapeutic doses of citicoline — given orally, by intramuscular injection, or by intravenous infusion — resulted in improvements in neuropsychological functions, including cognitive, emotional, and behavioral functions (reviewed in 5).
A six-month, multicenter observational study enrolled 197 stroke subjects (mean age, 81.5 years) with a progressive decline of their mental health and general confusion and/or stupor who were initially administered citicoline for 5 or 10 days (2,000 mg/day, by intravenous infusion) within a four-month period, and then for 21 days (1,000 mg/day, by intramuscular injection), repeated once after a seven-day washout period (93). Citicoline treatment was found to be associated with higher scores on cognitive and functional evaluation scales when compared to baseline measurements. However, only randomized controlled trials would be able to assess whether citicoline is protective against vascular damage and cognitive impairment in elderly adults with complex geriatric symptoms.
The International Citicoline Trial on acUte Stroke (ICTUS) is a multicenter and double-blind study that assessed the effect of supplementing 2,298 patients with acute ischemic stroke with citicoline (2,000 mg/day) or a placebo for six weeks on several functional and neurologic outcomes and on mortality rate (94). The results showed no difference between treatment groups after a 90-day follow-up period. Only subgroup analyses found significant benefits of citicoline in patients older than 70 years, in those with moderate rather than severe strokes, and in those not treated with recombinant tissue plasminogen activator (rtPA; standard-of-care treatment). An earlier meta-analysis of small randomized, placebo-controlled trials had reported a positive impact of citicoline (1,000 mg/day, administered for 28 days to 12 months) on memory and behavior in subjects with cognitive deficits associated with cerebrovascular disorders (95). The effect of citicoline was also evaluated in a multicenter, open-label, controlled trial (IDEALE trial) in Italian elderly adults (ages, 65-94 years) with evidence of vascular lesions on neuroradiology and mild-to-moderate cognitive deficits, as assessed by Mini-Mental State Examination (MMSE; scores ≥21) (96). Three hundred and forty-nine participants received oral citicoline (1,000 mg/day) or no treatment for nine months. MMSE scores in citicoline-treated individuals remained unchanged while they significantly deteriorated in untreated patients such that MMSE scores between groups were found to be significantly different after three and nine months of treatment. No significant effect was reported in measures of functional autonomy, mood, and behavioral disorders. Another open-label, randomized, controlled trial evaluated the effect of citicoline (1,000 mg/day for 12 months) in 347 subjects (mean age, 67.2 years) who suffered an acute stroke. The results demonstrated that citicoline significantly limited cognitive impairments in the domains of attention and executive functions and temporal orientation at 6 and 12 months post-stroke in treated compared to untreated patients (97). However, other randomized controlled trials have not found any benefit of citicoline treatment in the management of acute ischemic stroke (98, 99), including a recent trial evaluating its effects when provided immediately following endovascular thrombectomy (i.e., recanalization therapy) and continued for 42 days (100). A systematic review and meta-analysis of 10 randomized controlled trials found that citicoline therapy in acute ischemic stroke (provided intravenously and/or orally between 8 hours and 14 days post stroke) was linked to a slightly higher rate of independence (101). However, a recent Cochrane review of randomized controlled trials found that citicoline had no benefit to acute ischemic stroke patients compared to placebo with respect to all-cause mortality (8 trials); serious, adverse cardiovascular events (3 trials); or degree of disability or dependence with daily tasks (4 trials) (102).
Thus, there is little evidence that citicoline treatment is efficacious for patients with cerebrovascular disease. Many of the conducted trials have not been a double-blind design, which reduces the risk of bias.
For decades, preclinical and small clinical studies have investigated the effect of citicoline — when provided intravenously, intramuscularly, or orally — in the management of traumatic brain injury (TBI). A 2011 systematic review of clinical data suggested that citicoline could hasten the resorption of cerebral edema and improve the recovery of consciousness and neurologic disorders in severe TBI cases (classified by Glasgow Coma Scale [GCS] scores of ≤8) (5). Citicoline also appeared to limit memory deficits and the duration and severity of other post-traumatic symptoms (e.g., headache, dizziness, attention disorder) in TBI patients with mild-to-moderate injuries (GCS scores, 9-15) (reviewed in 5).
Although citicoline is included in TBI therapeutic regimen in 59 countries, only one multicenter, randomized, double-blind, placebo-controlled trial has been conducted in the US. The CiticOline Brain Injury Trial (COBRIT) has enrolled 1,213 patients with mild-to-severe TBI and assessed the effect of enteral or oral citicoline (2,000 mg/day, for 90 days) on functional and cognitive outcomes (measured by components of the TBI Clinical Trials Network Care Battery) (103). No significant benefits of citicoline supplementation over placebo were found at 90 days (end of treatment period) and 180 days. It is important to note that this trial had low adherence: only 44% of patients in the trial took at least 75% of their assigned dose (103). When pooling the results of COBRIT with those of 10 other clinical trials in a meta-analysis, citicoline treatment for acute-phase TBI was linked to a higher degree of independence, i.e., the capability of performing daily activities without needing assistance (104).
Humans can synthesize choline moieties in small amounts by converting phosphatidylethanolamine into phosphatidylcholine (Figure 2). Three methylation reactions catalyzed by phosphatidylethanolamine N-methyltransferase (PEMT) are required, each using S-adenosylmethionine (SAM) as a methyl group donor. Choline is generated endogenously when the methylation of phosphatidylethanolamine is coupled with the catabolism of newly formed phosphatidylcholine by phospholipases. This is referred to as de novo synthesis of choline. The substitution of choline by serine in the synthesis of phosphatidylserine from phosphatidylcholine by phosphatidylserine synthase-1 also releases choline (4). Because phosphatidylcholine metabolism is a source of endogenous choline, the nutrient was not initially classified as essential (1). Yet, de novo choline synthesis in humans is not sufficient to meet their metabolic needs such that healthy humans fed choline-deficient diets develop fatty liver, liver damage, and/or muscle damage (see Deficiency).
In the US, mean dietary intakes of choline are well below the recommended Adequate Intake (AI) level. According to a US national survey, NHANES 2015-2018, mean dietary intakes of choline were 284 mg/day for women and 390 mg/day for men; only 6% of women and 11% of men had dietary intakes greater than the AI (105). Moreover, NHANES data indicate that less than 9% of pregnant women meet the AI for choline (106). Americans of all ages have low intakes of choline: an analysis of NHANES 2009-2012 found that less than 11% of US residents ages 2 years and older had total choline intakes (from diet and supplements combined) above the AI (107). Stratifying the data by life stage indicated that young children ages 2-8 years were more likely to meet the AI than older children, adolescents, or adults (107). Vegetarians, especially vegans, who consume no meat, milk, or eggs, may be at risk for inadequate choline intake (26).
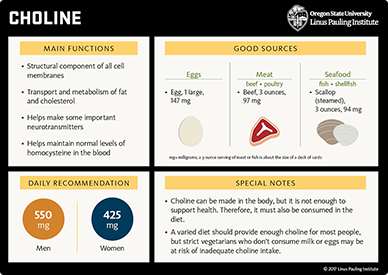
Eggs, liver, cruciferous vegetables, and peanuts are especially rich in choline (32, 108); one analysis of NHANES data concluded that it is extremely difficult to meet the AI from food sources alone if eggs are not consumed (106). Major contributors to choline in the American diet are meat, poultry, fish, dairy foods, pasta, rice, and egg-based dishes (109). Spinach, beets, wheat, and shellfish are also good sources of the choline metabolite, betaine (110). Betaine cannot be converted back to choline but can spare some choline requirements for homocysteine remethylation (1).
Phosphatidylcholine, which contains about 13% choline by weight, is the main form of choline in dietary products (111). Lecithin extracts, which comprise a mixture of phosphatidylcholine and other phospholipids, are often added during food processing. Lecithins in processed food have been estimated to increase the daily consumption of phosphatidylcholine by about 1.5 mg/kg of body weight for adults (32).
The total choline contents of some choline-containing foods are listed in milligrams (mg) in Table 2. For more information on the nutrient content of specific foods, search USDA’s FoodData Central or the USDA’s documentation on the choline content of common foods.
CDP-choline (citicoline) and choline salts, such as choline chloride and choline bitartrate, are available as supplements. Phosphatidylcholine supplements also provide choline; however, choline comprises only about 13% of the weight of phosphatidylcholine (111). Therefore, a supplement containing 4,230 mg (4.23 grams) of phosphatidylcholine would provide 550 mg of choline. Although the term "lecithin" is synonymous with phosphatidylcholine when used in chemistry, commercial lecithins are usually prepared from soybean, sunflower, and rapeseed, and may contain anywhere from 20%-90% of phosphatidylcholine. Egg yolk lecithin is a more unlikely source of lecithin in dietary supplements. Moreover, the nature of phosphatidylcholine-containing fatty acids depends on whether lecithin is produced from vegetable, animal, or microbial sources. In particular, soybean lecithin is richer in polyunsaturated fatty acids than egg yolk lecithin (112).
Most multivitamin supplements, including prenatal multivitamins, do not contain choline; the few that do often contain choline at levels much lower than the AI (113).
High doses (10,000 to 16,000 mg/day) of choline have been associated with a fishy body odor, vomiting, salivation, and increased sweating. The fishy body odor results from excessive production and excretion of trimethylamine, a metabolite of choline. In the inherited condition, primary trimethylaminuria (also known as "fish odor syndrome"; see the article on Riboflavin), a defective flavin containing monooxygenase 3 (FMO3) enzyme results in impaired oxidation of trimethylamine in the liver. Disease management includes the use of choline-restricted diets in affected individuals (114). Taking large doses of choline in the form of phosphatidylcholine (lecithin) does not generally result in fishy body odor, because its metabolism results in little trimethylamine.
A dose of 7,500 mg/day of choline was found to have a slight blood pressure-lowering (hypotensive) effect, which could result in dizziness or fainting. Choline magnesium trisalicylate at doses of 3,000 mg/day has resulted in impaired liver function, generalized itching, and ringing of the ears (tinnitus). However, it is likely that these effects were caused by the salicylate, rather than the choline in the preparation (32).
In 1998, the Food and Nutrition Board (FNB) of the Institute of Medicine (now the National Academy of Medicine) established the tolerable upper intake level (UL) for choline at 3,500 mg/day for adults (Table 3). This recommendation was based primarily on preventing hypotension (low blood pressure), and secondarily, on preventing the fishy body odor due to increased excretion of trimethylamine. The UL was established for generally healthy people, and the FNB noted that individuals with liver or kidney disease, Parkinson's disease, depression, or inherited trimethylaminuria might be at increased risk of adverse effects when consuming choline at levels near the UL (32).
Oral supplementation with phosphatidylcholine (250 mg of total choline from food plus 250 mg of supplemental phosphatidylcholine) has been found to result in detectable concentrations of trimethylamine and trimethylamine N-oxide (TMAO) in the blood (23). The intestinal microbiota is directly implicated in the generation of trimethylamine from dietary choline and its metabolite phosphatidylcholine, as well as from dietary betaine and carnitine. Trimethylamine is subsequently converted into TMAO by flavin-containing monooxygenases in the liver. The prospective study that followed 4,007 individuals — with or without cardiovascular disease — for a three-year period found baseline concentrations of circulating TMAO to be positively correlated with incidence of death, nonfatal myocardial infarction, and stroke — described as major adverse cardiac events (MACE) (23). In the same cohort, MACE risk was found to be about 30% higher in individuals in the highest versus lowest quartile of choline or betaine plasma concentrations (115). However, depending on gut microbiota composition, the risk of having an adverse cardiovascular event may be lower in individuals with low versus high circulating TMAO even though choline and/or betaine concentrations in the blood are elevated (115). Similar findings have since been made in other prospective cohorts. A meta-analysis of 19 prospective studies found that higher blood TMAO concentrations were associated with a 62% increased risk for MACE or death compared to lower concentrations (116). Elevated circulating TMAO concentrations have been not only linked to an increased risk of cardiovascular disease but also to an increased risk for type 2 diabetes mellitus and kidney disease (117-119). However, high TMAO concentrations in blood may be a biomarker of the disease rather than a causative factor (120, 121).
Further research is needed to understand how the composition of intestinal microbiota influences the metabolic fate of ingested choline. At present, there is little evidence that dietary choline increases the risk of cardiovascular events. Prospective cohort studies on the association have been inconsistent: a few studies have linked higher choline intakes to increased risks of atrial fibrillation (122) and cardiovascular disease-related mortality (43, 44), while several other prospective studies have found no association between choline intake and cardiovascular disease endpoints (41, 45-48). Interestingly, in the PREvención con DIeta MEDiterránea-Plus (Predimed-Plus) trial in overweight or obese individuals with metabolic syndrome, an increase in dietary choline intake over a one-year period was associated with improved measures of cardiovascular health (i.e., lower serum total cholesterol, serum LDL cholesterol, and systolic and diastolic blood pressure levels) (123).
Methotrexate, a medication used in the treatment of cancer, psoriasis, and rheumatoid arthritis, inhibits the enzyme dihydrofolate reductase and therefore limits the availability of methyl groups donated from folate derivatives. Rats given methotrexate have shown evidence of diminished nutritional status of choline and greater drug adverse reactions due to liver dysfunction (12, 124). Thus, individuals taking methotrexate may have an increased choline requirement. Treatments with a family of lipid-lowering drugs known as fibrates (e.g., fenofibrate, bezofibrate) have been associated with an increased excretion of betaine in the urine and a rise in homocysteine concentration in the blood of patients with diabetes mellitus or metabolic syndrome (125, 126). If the benefits of fibrate therapy are indeed mitigated by fibrate-induced betaine deficiency, the use and safety of supplementing patients with betaine would need to be considered (127).
Little is known regarding the amount of dietary choline required to promote optimum health or prevent chronic diseases in humans. The Linus Pauling Institute supports the recommendation by the Food and Nutrition Board of 425 mg/day for adult women and 550 mg/day for adult men. A varied, healthy diet should provide enough choline for most people, but strict vegetarians who consume no milk or eggs may be at risk of inadequate choline intake.
Little is known regarding the amount of dietary choline most likely to promote optimum health or prevent chronic diseases in older adults. At present, there is no evidence to support a different recommended intake of choline from that of younger adults (425 mg/day for women and 550 mg/day for men).
Originally written in 2000 by:
Jane Higdon, Ph.D.
Linus Pauling Institute
Oregon State University
Updated in May 2003 by:
Jane Higdon, Ph.D.
Linus Pauling Institute
Oregon State University
Updated in January 2008 by:
Victoria J. Drake, Ph.D.
Linus Pauling Institute
Oregon State University
Updated in August 2009 by:
Victoria J. Drake, Ph.D.
Linus Pauling Institute
Oregon State University
Updated in January 2015 by:
Barbara Delage, Ph.D.
Linus Pauling Institute
Oregon State University
Updated in April 2023 by:
Victoria J. Drake, Ph.D.
Linus Pauling Institute
Oregon State University
Reviewed in July 2023 by:
Isis Trujillo-Gonzalez, Ph.D.
Assistant Professor
Nutrition Research Institute
University of North Carolina at Chapel Hill
Copyright 2000-2024 Linus Pauling Institute
1. Zeisel SH, Corbin KD. Choline. Present Knowledge in Nutrition. 10th ed: John Wiley & Sons, Inc.; 2012:405-418.
2. Ueland PM. Choline and betaine in health and disease. J Inherit Metab Dis. 2011;34(1):3-15. (PubMed)
3. Gibellini F, Smith TK. The Kennedy pathway--De novo synthesis of phosphatidylethanolamine and phosphatidylcholine. IUBMB Life. 2010;62(6):414-428. (PubMed)
4. Li Z, Vance DE. Phosphatidylcholine and choline homeostasis. J Lipid Res. 2008;49(6):1187-1194. (PubMed)
5. Secades JJ. Citicoline: pharmacological and clinical review, 2010 update. Rev Neurol. 2011;52 Suppl 2:S1-S62. (PubMed)
6. Beckmann J, Lips KS. The non-neuronal cholinergic system in health and disease. Pharmacology. 2013;92(5-6):286-302. (PubMed)
7. Noga AA, Vance DE. A gender-specific role for phosphatidylethanolamine N-methyltransferase-derived phosphatidylcholine in the regulation of plasma high density and very low density lipoproteins in mice. J Biol Chem. 2003;278(24):21851-21859. (PubMed)
8. Noga AA, Zhao Y, Vance DE. An unexpected requirement for phosphatidylethanolamine N-methyltransferase in the secretion of very low density lipoproteins. J Biol Chem. 2002;277(44):42358-42365. (PubMed)
9. da Costa KA, Kozyreva OG, Song J, Galanko JA, Fischer LM, Zeisel SH. Common genetic polymorphisms affect the human requirement for the nutrient choline. Faseb J. 2006;20(9):1336-1344. (PubMed)
10. Pellanda H. Betaine homocysteine methyltransferase (BHMT)-dependent remethylation pathway in human healthy and tumoral liver. Clin Chem Lab Med. 2013;51(3):617-621. (PubMed)
11. Gerhard GT, Duell PB. Homocysteine and atherosclerosis. Curr Opin Lipidol. 1999;10(5):417-428. (PubMed)
12. Zeisel SH. Choline. In: Ross A, Caballero B, Cousins R, Tucker K, Ziegler T, eds. Modern Nutrition in Health and Disease. 11th ed: Lippincott Williams & Wilkins; 2014:416-426.
13. Michelotti GA, Machado MV, Diehl AM. NAFLD, NASH and liver cancer. Nat Rev Gastroenterol Hepatol. 2013;10(11):656-665. (PubMed)
14. Zeisel SH, Blusztajn JK. Choline and human nutrition. Annu Rev Nutr. 1994;14:269-296. (PubMed)
15. da Costa KA, Niculescu MD, Craciunescu CN, Fischer LM, Zeisel SH. Choline deficiency increases lymphocyte apoptosis and DNA damage in humans. Am J Clin Nutr. 2006;84(1):88-94. (PubMed)
16. Rolo AP, Teodoro JS, Palmeira CM. Role of oxidative stress in the pathogenesis of nonalcoholic steatohepatitis. Free Radic Biol Med. 2012;52(1):59-69. (PubMed)
17. Fischer LM, daCosta KA, Kwock L, et al. Sex and menopausal status influence human dietary requirements for the nutrient choline. Am J Clin Nutr. 2007;85(5):1275-1285. (PubMed)
18. Fischer LM, da Costa KA, Kwock L, Galanko J, Zeisel SH. Dietary choline requirements of women: effects of estrogen and genetic variation. Am J Clin Nutr. 2010;92(5):1113-1119. (PubMed)
19. Resseguie M, Song J, Niculescu MD, da Costa KA, Randall TA, Zeisel SH. Phosphatidylethanolamine N-methyltransferase (PEMT) gene expression is induced by estrogen in human and mouse primary hepatocytes. Faseb J. 2007;21(10):2622-2632. (PubMed)
20. da Costa KA, Corbin KD, Niculescu MD, Galanko JA, Zeisel SH. Identification of new genetic polymorphisms that alter the dietary requirement for choline and vary in their distribution across ethnic and racial groups. FASEB J. 2014;28(7):2970-2978. (PubMed)
21. Kohlmeier M, da Costa KA, Fischer LM, Zeisel SH. Genetic variation of folate-mediated one-carbon transfer pathway predicts susceptibility to choline deficiency in humans. Proc Natl Acad Sci U S A. 2005;102(44):16025-16030. (PubMed)
22. Spencer MD, Hamp TJ, Reid RW, Fischer LM, Zeisel SH, Fodor AA. Association between composition of the human gastrointestinal microbiome and development of fatty liver with choline deficiency. Gastroenterology. 2011;140(3):976-986. (PubMed)
23. Tang WH, Wang Z, Levison BS, et al. Intestinal microbial metabolism of phosphatidylcholine and cardiovascular risk. N Engl J Med. 2013;368(17):1575-1584. (PubMed)
24. Wang Z, Klipfell E, Bennett BJ, et al. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature. 2011;472(7341):57-63. (PubMed)
25. Korsmo HW, Jiang X, Caudill MA. Choline: exploring the growing science on its benefits for moms and babies. Nutrients. 2019;11(8). (PubMed)
26. Wallace TC, Blusztajn JK, Caudill MA, et al. Choline: the underconsumed and underappreciated essential nutrient. Nutr Today. 2018;53(6):240-253. (PubMed)
27. Bernhard W, Shunova A, Machann J, et al. Resolution of severe hepatosteatosis in a cystic fibrosis patient with multifactorial choline deficiency: A case report. Nutrition. 2021;89:111348. (PubMed)
28. Grothe J, Riethmuller J, Tschurtz SM, et al. Plasma phosphatidylcholine alterations in cystic fibrosis patients: impaired metabolism and correlation with lung function and inflammation. Cell Physiol Biochem. 2015;35(4):1437-1453. (PubMed)
29. Bernhard W, Poets CF, Franz AR. Choline and choline-related nutrients in regular and preterm infant growth. Eur J Nutr. 2019;58(3):931-945. (PubMed)
30. Jacob RA, Jenden DJ, Allman-Farinelli MA, Swendseid ME. Folate nutriture alters choline status of women and men fed low choline diets. J Nutr. 1999;129(3):712-717. (PubMed)
31. Kim YI, Miller JW, da Costa KA, et al. Severe folate deficiency causes secondary depletion of choline and phosphocholine in rat liver. J Nutr. 1994;124(11):2197-2203. (PubMed)
32. Food and Nutrition Board, Institute of Medicine. Choline. Dietary Reference Intakes: Thiamin, Riboflavin, Niacin, Vitamin B-6, Vitamin B-12, Pantothenic Acid, Biotin, and Choline. Washington D.C.: National Academy Press; 1998:390-422. (National Academy Press)
33. Ganz AB, Klatt KC, Caudill MA. Common genetic variants alter metabolism and influence dietary choline requirements. Nutrients. 2017;9(8):252. (PubMed)
34. Ganz AB, Cohen VV, Swersky CC, et al. Genetic variation in choline-metabolizing enzymes alters choline metabolism in young women consuming choline intakes meeting current recommendations. Int J Mol Sci. 2017;18(2):837. (PubMed)
35. Zhou J, Austin RC. Contributions of hyperhomocysteinemia to atherosclerosis: Causal relationship and potential mechanisms. Biofactors. 2009;35(2):120-129. (PubMed)
36. Leach NV, Dronca E, Vesa SC, et al. Serum homocysteine levels, oxidative stress and cardiovascular risk in non-alcoholic steatohepatitis. Eur J Intern Med. 2014;25(8):762-767. (PubMed)
37. Olthof MR, Brink EJ, Katan MB, Verhoef P. Choline supplemented as phosphatidylcholine decreases fasting and postmethionine-loading plasma homocysteine concentrations in healthy men. Am J Clin Nutr. 2005;82(1):111-117. (PubMed)
38. Olthof MR, van Vliet T, Boelsma E, Verhoef P. Low dose betaine supplementation leads to immediate and long term lowering of plasma homocysteine in healthy men and women. J Nutr. 2003;133(12):4135-4138. (PubMed)
39. Schwab U, Torronen A, Toppinen L, et al. Betaine supplementation decreases plasma homocysteine concentrations but does not affect body weight, body composition, or resting energy expenditure in human subjects. Am J Clin Nutr. 2002;76(5):961-967. (PubMed)
40. Steenge GR, Verhoef P, Katan MB. Betaine supplementation lowers plasma homocysteine in healthy men and women. J Nutr. 2003;133(5):1291-1295. (PubMed)
41. Dalmeijer GW, Olthof MR, Verhoef P, Bots ML, van der Schouw YT. Prospective study on dietary intakes of folate, betaine, and choline and cardiovascular disease risk in women. Eur J Clin Nutr. 2008;62(3):386-394. (PubMed)
42. Millard HR, Musani SK, Dibaba DT, et al. Dietary choline and betaine; associations with subclinical markers of cardiovascular disease risk and incidence of CVD, coronary heart disease and stroke: the Jackson Heart Study. Eur J Nutr. 2018;57(1):51-60. (PubMed)
43. Zheng Y, Li Y, Rimm EB, et al. Dietary phosphatidylcholine and risk of all-cause and cardiovascular-specific mortality among US women and men. Am J Clin Nutr. 2016;104(1):173-180. (PubMed)
44. Yang JJ, Lipworth LP, Shu XO, et al. Associations of choline-related nutrients with cardiometabolic and all-cause mortality: results from 3 prospective cohort studies of blacks, whites, and Chinese. Am J Clin Nutr. 2020;111(3):644-656. (PubMed)
45. Nagata C, Wada K, Tamura T, et al. Choline and betaine intakes are not associated with cardiovascular disease mortality risk in Japanese men and women. J Nutr. 2015;145(8):1787-1792. (PubMed)
46. Bidulescu A, Chambless LE, Siega-Riz AM, Zeisel SH, Heiss G. Usual choline and betaine dietary intake and incident coronary heart disease: the Atherosclerosis Risk in Communities (ARIC) study. BMC Cardiovasc Disord. 2007;7:20. (PubMed)
47. Bertoia ML, Pai JK, Cooke JP, et al. Plasma homocysteine, dietary B vitamins, betaine, and choline and risk of peripheral artery disease. Atherosclerosis. 2014;235(1):94-101. (PubMed)
48. Golzarand M, Mirmiran P, Azizi F. Association between dietary choline and betaine intake and 10.6-year cardiovascular disease in adults. Nutr J. 2022;21(1):1. (PubMed)
49. Lundberg P, Dudman NP, Kuchel PW, Wilcken DE. 1H NMR determination of urinary betaine in patients with premature vascular disease and mild homocysteinemia. Clin Chem. 1995;41(2):275-283. (PubMed)
50. Lever M, George PM, Elmslie JL, et al. Betaine and secondary events in an acute coronary syndrome cohort. PLoS One. 2012;7(5):e37883. (PubMed)
51. Konstantinova SV, Tell GS, Vollset SE, Nygard O, Bleie O, Ueland PM. Divergent associations of plasma choline and betaine with components of metabolic syndrome in middle age and elderly men and women. J Nutr. 2008;138(5):914-920. (PubMed)
52. Meyer KA, Benton TZ, Bennett BJ, et al. Microbiota-dependent metabolite trimethylamine N-oxide and coronary artery calcium in the coronary artery risk development in young adults study (CARDIA). J Am Heart Assoc. 2016;5(10). (PubMed)
53. Yu D, Shu XO, Xiang YB, et al. Higher dietary choline intake is associated with lower risk of nonalcoholic fatty liver in normal-weight Chinese women. J Nutr. 2014;144(12):2034-2040. (PubMed)
54. Guerrerio AL, Colvin RM, Schwartz AK, et al. Choline intake in a large cohort of patients with nonalcoholic fatty liver disease. Am J Clin Nutr. 2012;95(4):892-900. (PubMed)
55. Mazidi M, Katsiki N, Mikhailidis DP, Banach M. Adiposity may moderate the link between choline intake and non-alcoholic fatty liver disease. J Am Coll Nutr. 2019;38(7):633-639. (PubMed)
56. Talaulikar VS, Arulkumaran S. Folic acid in obstetric practice: a review. Obstet Gynecol Surv. 2011;66(4):240-247. (PubMed)
57. Czeizel AE, Dudas I, Vereczkey A, Banhidy F. Folate deficiency and folic acid supplementation: the prevention of neural-tube defects and congenital heart defects. Nutrients. 2013;5(11):4760-4775. (PubMed)
58. Eskes TK. Open or closed? A world of difference: a history of homocysteine research. Nutr Rev. 1998;56(8):236-244. (PubMed)
59. Shaw GM, Carmichael SL, Yang W, Selvin S, Schaffer DM. Periconceptional dietary intake of choline and betaine and neural tube defects in offspring. Am J Epidemiol. 2004;160(2):102-109. (PubMed)
60. Carmichael SL, Yang W, Shaw GM. Periconceptional nutrient intakes and risks of neural tube defects in California. Birth Defects Res A Clin Mol Teratol. 2010;88(8):670-678. (PubMed)
61. Chandler AL, Hobbs CA, Mosley BS, et al. Neural tube defects and maternal intake of micronutrients related to one-carbon metabolism or antioxidant activity. Birth Defects Res A Clin Mol Teratol. 2012;94(11):864-874. (PubMed)
62. Petersen JM, Parker SE, Crider KS, Tinker SC, Mitchell AA, Werler MM. One-carbon cofactor intake and risk of neural tube defects among women who meet folic acid recommendations: a multicenter case-control study. Am J Epidemiol. 2019;188(6):1136-1143. (PubMed)
63. Lavery AM, Brender JD, Zhao H, et al. Dietary intake of choline and neural tube defects in Mexican Americans. Birth Defects Res A Clin Mol Teratol. 2014;100(6):463-471. (PubMed)
64. Shaw GM, Finnell RH, Blom HJ, et al. Choline and risk of neural tube defects in a folate-fortified population. Epidemiology. 2009;20(5):714-719. (PubMed)
65. Mills JL, Fan R, Brody LC, et al. Maternal choline concentrations during pregnancy and choline-related genetic variants as risk factors for neural tube defects. Am J Clin Nutr. 2014;100(4):1069-1074. (PubMed)
66. Obeid R, Derbyshire E, Schon C. Association between maternal choline, fetal brain development, and child neurocognition: systematic review and meta-analysis of human studies. Adv Nutr. 2022;13(6):2445-2457. (PubMed)
67. Teather LA, Wurtman RJ. Dietary cytidine (5')-diphosphocholine supplementation protects against development of memory deficits in aging rats. Prog Neuropsychopharmacol Biol Psychiatry. 2003;27(4):711-717. (PubMed)
68. Zeisel SH. Choline: critical role during fetal development and dietary requirements in adults. Annu Rev Nutr. 2006;26:229-250. (PubMed)
69. McCann JC, Hudes M, Ames BN. An overview of evidence for a causal relationship between dietary availability of choline during development and cognitive function in offspring. Neurosci Biobehav Rev. 2006;30(5):696-712. (PubMed)
70. Dauncey MJ. Nutrition, the brain and cognitive decline: insights from epigenetics. Eur J Clin Nutr. 2014;68(11):1179-1185. (PubMed)
71. Strain JJ, McSorley EM, van Wijngaarden E, et al. Choline status and neurodevelopmental outcomes at 5 years of age in the Seychelles Child Development Nutrition Study. Br J Nutr. 2013;110(2):330-336. (PubMed)
72. Villamor E, Rifas-Shiman SL, Gillman MW, Oken E. Maternal intake of methyl-donor nutrients and child cognition at 3 years of age. Paediatr Perinat Epidemiol. 2012;26(4):328-335. (PubMed)
73. Boeke CE, Gillman MW, Hughes MD, Rifas-Shiman SL, Villamor E, Oken E. Choline intake during pregnancy and child cognition at age 7 years. Am J Epidemiol. 2013;177(12):1338-1347. (PubMed)
74. Cheatham CL, Goldman BD, Fischer LM, da Costa KA, Reznick JS, Zeisel SH. Phosphatidylcholine supplementation in pregnant women consuming moderate-choline diets does not enhance infant cognitive function: a randomized, double-blind, placebo-controlled trial. Am J Clin Nutr. 2012;96(6):1465-1472. (PubMed)
75. Caudill MA, Strupp BJ, Muscalu L, Nevins JEH, Canfield RL. Maternal choline supplementation during the third trimester of pregnancy improves infant information processing speed: a randomized, double-blind, controlled feeding study. FASEB J. 2018;32(4):2172-2180. (PubMed)
76. Bahnfleth CL, Strupp BJ, Caudill MA, Canfield RL. Prenatal choline supplementation improves child sustained attention: A 7-year follow-up of a randomized controlled feeding trial. FASEB J. 2022;36(1):e22054. (PubMed)
77. Wang Z, Zhu W, Xing Y, Jia J, Tang Y. B vitamins and prevention of cognitive decline and incident dementia: a systematic review and meta-analysis. Nutr Rev. 2022;80(4):931-949. (PubMed)
78. Poly C, Massaro JM, Seshadri S, et al. The relation of dietary choline to cognitive performance and white-matter hyperintensity in the Framingham Offspring Cohort. Am J Clin Nutr. 2011;94(6):1584-1591. (PubMed)
79. Liu L, Qiao S, Zhuang L, et al. Choline intake correlates with cognitive performance among elder adults in the United States. Behav Neurol. 2021;2021:2962245. (PubMed)
80. Nurk E, Refsum H, Bjelland I, et al. Plasma free choline, betaine and cognitive performance: the Hordaland Health Study. Br J Nutr. 2013;109(3):511-519. (PubMed)
81. Eussen SJ, Ueland PM, Clarke R, et al. The association of betaine, homocysteine and related metabolites with cognitive function in Dutch elderly people. Br J Nutr. 2007;98(5):960-968. (PubMed)
82. Birks J. Cholinesterase inhibitors for Alzheimer's disease. Cochrane Database Syst Rev. 2006(1):CD005593. (PubMed)
83. Higgins JP, Flicker L. Lecithin for dementia and cognitive impairment. Cochrane Database Syst Rev. 2003(3):CD001015. (PubMed)
84. Bonvicini M, Travaglini S, Lelli D, Antonelli Incalzi R, Pedone C. Is citicoline effective in preventing and slowing down dementia?-a systematic review and a meta-analysis. Nutrients. 2023;15(2):386. (PubMed)
85. Gupta N, Yucel YH. Glaucoma as a neurodegenerative disease. Curr Opin Ophthalmol. 2007;18(2):110-114. (PubMed)
86. Hwang JS, Shin YJ. Role of choline in ocular diseases. Int J Mol Sci. 2021;22(9):4733. (PubMed)
87. Parisi V. Electrophysiological assessment of glaucomatous visual dysfunction during treatment with cytidine-5'-diphosphocholine (citicoline): a study of 8 years of follow-up. Doc Ophthalmol. 2005;110(1):91-102. (PubMed)
88. Parisi V, Coppola G, Ziccardi L, Gallinaro G, Falsini B. Cytidine-5'-diphosphocholine (Citicoline): a pilot study in patients with non-arteritic ischaemic optic neuropathy. Eur J Neurol. 2008;15(5):465-474. (PubMed)
89. Ottobelli L, Manni GL, Centofanti M, Iester M, Allevena F, Rossetti L. Citicoline oral solution in glaucoma: is there a role in slowing disease progression? Ophthalmologica. 2013;229(4):219-226. (PubMed)
90. Parisi V, Centofanti M, Ziccardi L, et al. Treatment with citicoline eye drops enhances retinal function and neural conduction along the visual pathways in open angle glaucoma. Graefes Arch Clin Exp Ophthalmol. 2015;253(8):1327-1340. (PubMed)
91. Rossetti L, Iester M, Tranchina L, et al. Can treatment with citicoline eyedrops reduce progression in glaucoma? The results of a randomized placebo-controlled clinical trial. J Glaucoma. 2020;29(7):513-520. (PubMed)
92. Grieb P. Neuroprotective properties of citicoline: facts, doubts and unresolved issues. CNS Drugs. 2014;28(3):185-193. (PubMed)
93. Putignano S, Gareri P, Castagna A, et al. Retrospective and observational study to assess the efficacy of citicoline in elderly patients suffering from stupor related to complex geriatric syndrome. Clin Interv Aging. 2012;7:113-118. (PubMed)
94. Davalos A, Alvarez-Sabin J, Castillo J, et al. Citicoline in the treatment of acute ischaemic stroke: an international, randomised, multicentre, placebo-controlled study (ICTUS trial). Lancet. 2012;380(9839):349-357. (PubMed)
95. Fioravanti M, Yanagi M. Cytidinediphosphocholine (CDP-choline) for cognitive and behavioural disturbances associated with chronic cerebral disorders in the elderly. Cochrane Database Syst Rev. 2005(2):CD000269. (PubMed)
96. Cotroneo AM, Castagna A, Putignano S, et al. Effectiveness and safety of citicoline in mild vascular cognitive impairment: the IDEALE study. Clin Interv Aging. 2013;8:131-137. (PubMed)
97. Alvarez-Sabin J, Ortega G, Jacas C, et al. Long-term treatment with citicoline may improve poststroke vascular cognitive impairment. Cerebrovasc Dis. 2013;35(2):146-154. (PubMed)
98. Clark WM, Williams BJ, Selzer KA, Zweifler RM, Sabounjian LA, Gammans RE. A randomized efficacy trial of citicoline in patients with acute ischemic stroke. Stroke. 1999;30(12):2592-2597. (PubMed)
99. Clark WM, Wechsler LR, Sabounjian LA, Schwiderski UE, Citicoline Stroke Study Group. A phase III randomized efficacy trial of 2000 mg citicoline in acute ischemic stroke patients. Neurology. 2001;57(9):1595-1602. (PubMed)
100. Agarwal A, Vishnu VY, Sharma J, et al. Citicoline in acute ischemic stroke: A randomized controlled trial. PLoS One. 2022;17(5):e0269224. (PubMed)
101. Secades JJ, Alvarez-Sabin J, Castillo J, et al. Citicoline for acute ischemic atroke: a systematic review and formal meta-analysis of randomized, double-blind, and placebo-controlled trials. J Stroke Cerebrovasc Dis. 2016;25(8):1984-1996. (PubMed)
102. Marti-Carvajal AJ, Valli C, Marti-Amarista CE, Sola I, Marti-Fabregas J, Bonfill Cosp X. Citicoline for treating people with acute ischemic stroke. Cochrane Database Syst Rev. 2020;8(8):CD013066. (PubMed)
103. Zafonte RD, Bagiella E, Ansel BM, et al. Effect of citicoline on functional and cognitive status among patients with traumatic brain injury: Citicoline Brain Injury Treatment Trial (COBRIT). JAMA. 2012;308(19):1993-2000. (PubMed)
104. Secades JJ, Trimmel H, Salazar B, Gonzalez JA. Citicoline for the management of patients with traumatic brain injury in the acute phase: a systematic review and meta-analysis. Life (Basel). 2023;13(2):369. (PubMed)
105. USDA, Agricultural Research Service, 2021. Usual Nutrient Intake from Food and Beverages, by Gender and Age, What We Eat in America, NHANES 2015-2018. Available http://www.ars.usda.gov/nea/bhnrc/fsrg.
106. Wallace TC, Fulgoni VL 3rd. Usual choline intakes are associated with egg and protein food consumption in the United States. Nutrients. 2017;9(8):839. (PubMed)
107. Wallace TC, Fulgoni VL, 3rd. Assessment of total choline intakes in the United States. J Am Coll Nutr. 2016;35(2):108-112. (PubMed)
108. Zeisel SH, Klatt KC, Caudill MA. Choline. Adv Nutr. 2018;9(1):58-60. (PubMed)
109. Chester DN, Goldman JD, Ahuja JK, Moshfegh AJ. Dietary Intakes of Choline: What We Eat in America, NHANES 2007-2008. Food Surveys Research Group Dietary Data Brief No. 9. October 2011. Available at: http://ars.usda.gov/Services/docs.htm?docid=19476.
110. Craig SA. Betaine in human nutrition. Am J Clin Nutr. 2004;80(3):539-549. (PubMed)
111. Koc H, Mar MH, Ranasinghe A, Swenberg JA, Zeisel SH. Quantitation of choline and its metabolites in tissues and foods by liquid chromatography/electrospray ionization-isotope dilution mass spectrometry. Anal Chem. 2002;74(18):4734-4740. (PubMed)
112. Hendler SS, Rorvik DR, eds. PDR for Nutritional Supplements. 2nd ed: Thomson Reuters; 2008.
113. US Department of Health and Human Services, National Institutes of Health, Office of Dietary Supplements. Dietary Supplement Label Database (DSLD). [Internet]. [cited 4/10/2023]. Available from: https://dsld.od.nih.gov/.
114. Busby MG, Fischer L, da Costa KA, Thompson D, Mar MH, Zeisel SH. Choline- and betaine-defined diets for use in clinical research and for the management of trimethylaminuria. J Am Diet Assoc. 2004;104(12):1836-1845. (PubMed)
115. Wang Z, Tang WH, Buffa JA, et al. Prognostic value of choline and betaine depends on intestinal microbiota-generated metabolite trimethylamine-N-oxide. Eur Heart J. 2014;35(14):904-910. (PubMed)
116. Heianza Y, Ma W, Manson JE, Rexrode KM, Qi L. Gut microbiota metabolites and risk of major adverse cardiovascular disease events and death: a systematic review and meta-analysis of prospective studies. J Am Heart Assoc. 2017;6(7):e004947. (PubMed)
117. Papandreou C, More M, Bellamine A. Trimethylamine N-oxide in relation to cardiometabolic health-cause or effect? Nutrients. 2020;12(5):1330. (PubMed)
118. Zeisel SH, Warrier M. Trimethylamine N-oxide, the microbiome, and heart and kidney disease. Annu Rev Nutr. 2017;37:157-181. (PubMed)
119. Gatarek P, Kaluzna-Czaplinska J. Trimethylamine N-oxide (TMAO) in human health. EXCLI J. 2021;20:301-319. (PubMed)
120. Canyelles M, Borras C, Rotllan N, Tondo M, Escola-Gil JC, Blanco-Vaca F. Gut microbiota-derived TMAO: a causal factor promoting atherosclerotic cardiovascular disease? Int J Mol Sci. 2023;24(3):1940. (PubMed)
121. Al-Obaide MAI, Singh R, Datta P, et al. Gut microbiota-dependent trimethylamine-N-oxide and serum biomarkers in patients with T2DM and advanced CKD. J Clin Med. 2017;6(9):86. (PubMed)
122. Zuo H, Svingen GFT, Tell GS, et al. Plasma concentrations and dietary intakes of choline and betaine in association with atrial fibrillation risk: results from 3 prospective cohorts with different health profiles. J Am Heart Assoc. 2018;7(8):e008190. (PubMed)
123. Diez-Ricote L, San-Cristobal R, Concejo MJ, et al. One-year longitudinal association between changes in dietary choline or betaine intake and cardiometabolic variables in the PREvencion con DIeta MEDiterranea-Plus (PREDIMED-Plus) trial. Am J Clin Nutr. 2022;116(6):1565-1579. (PubMed)
124. Hardwick RN, Clarke JD, Lake AD, et al. Increased susceptibility to methotrexate-induced toxicity in nonalcoholic steatohepatitis. Toxicol Sci. 2014;142(1):45-55. (PubMed)
125. Lever M, George PM, Slow S, et al. Fibrates may cause an abnormal urinary betaine loss which is associated with elevations in plasma homocysteine. Cardiovasc Drugs Ther. 2009;23(5):395-401. (PubMed)
126. Lever M, McEntyre CJ, George PM, et al. Extreme urinary betaine losses in type 2 diabetes combined with bezafibrate treatment are associated with losses of dimethylglycine and choline but not with increased losses of other osmolytes. Cardiovasc Drugs Ther. 2014;28(5):459-468. (PubMed)
127. Lever M, George PM, Slow S, et al. Fibrates plus betaine: a winning combination? N Z Med J. 2010;123(1324):74-78. (PubMed)