Summary
- Cruciferous vegetables are unique in that they are rich sources of sulfur-containing compounds known as glucosinolates. (More information)
- Chopping or chewing raw cruciferous vegetables results in the formation of bioactive glucosinolate hydrolysis products, such as isothiocyanates and indole-3-carbinol. These metabolites are also generated by colonic bacteria, following the ingestion of cooked cruciferous vegetables. (More information)
- The biological activities of glucosinolate-derived isothiocyanates and indole-3-carbinol likely contribute to the potential health-promoting effects of cruciferous vegetables. (More information)
- Variations in the sequence of genes coding for Phase II detoxification enzymes, glutathione S-transferases (GSTs), may influence the potential health benefits of consuming cruciferous vegetables. (More information)
- A few observational studies have examined the link between cruciferous vegetable consumption and cardiovascular health, but the results have been largely inconsistent. (More information)
- High intakes of cruciferous vegetables have been associated with lower risk of bladder, breast, colorectal, endometrial, gastric, lung, ovarian, pancreatic, prostate, and renal cancer. However, evidence of associations remains largely limited to case-control studies. (More information)
- The Dietary Guidelines for Americans recommend for adults to consume 1½-2½ cup-equivalents of dark-green vegetables per week, including cruciferous vegetables, as part of healthy meals. (More information)
Other Articles
Introduction
Cruciferous or Brassica vegetables come from plants in the family known to botanists and biologists as Cruciferae or alternately, Brassicaceae. The Brassicaceae family, which includes the model plant Arabidopsis thaliana, comprises approximately 375 genera and over 3,000 species (1). Many, but not all, commonly consumed cruciferous vegetables come from the Brassica genus; examples include broccoli, Brussels sprouts, cabbage, cauliflower, collard greens, kale, kohlrabi, mustard, rutabaga, turnips, bok choy, and Chinese cabbage (2). Examples of other edible crucifers include radish (Raphanus sativus), horseradish (Armoracia rusticana), watercress (Nasturtium officinale), and wasabi (Wasabia japonica) (2).
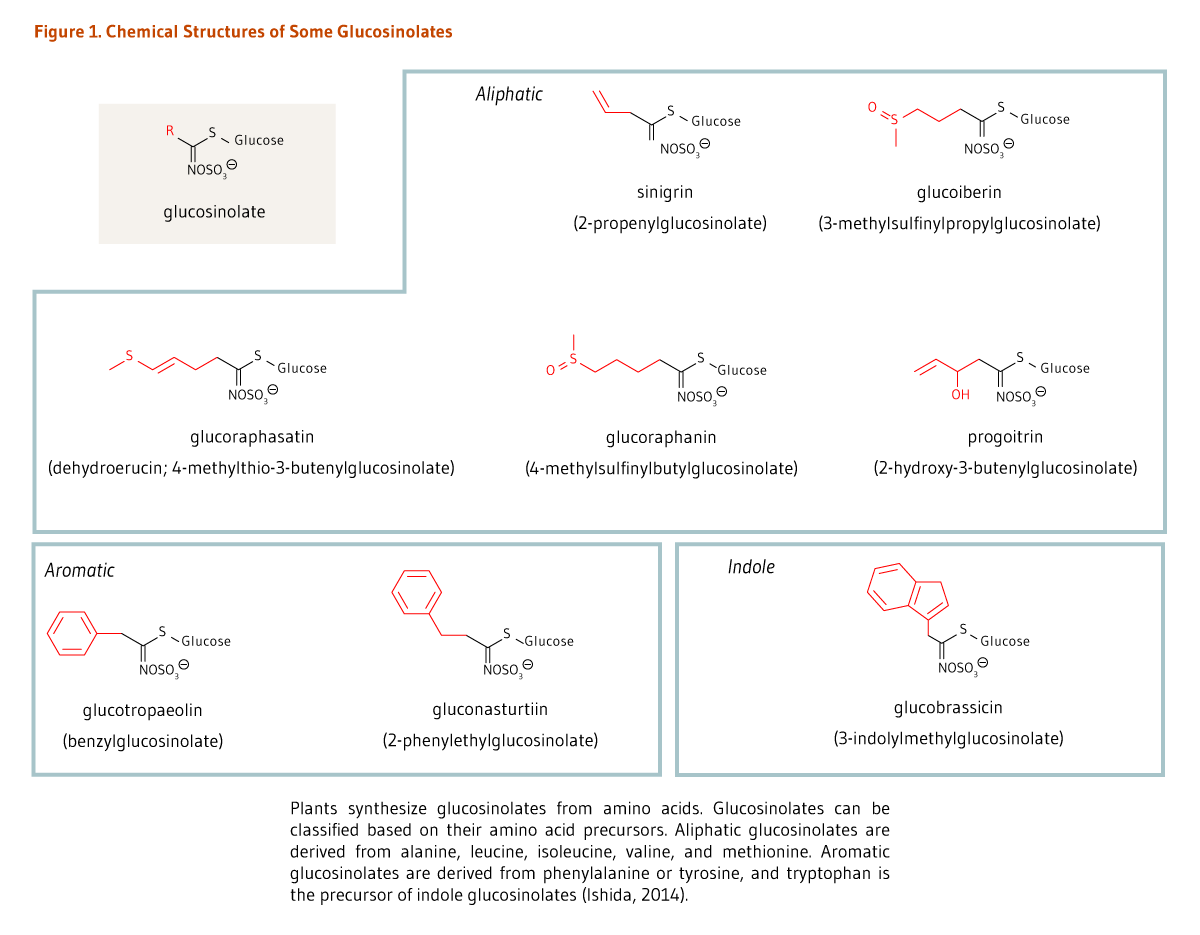
Cruciferous vegetables are unique in that they are a rich source of sulfur-containing compounds called glucosinolates (β-thioglucoside N-hydroxysulfates) that impart a pungent aroma and spicy (some say bitter) taste (Figure 1). Glucosinolates can be classified into three categories based on the chemical structure of their amino acid precursors: aliphatic glucosinolates (e.g., glucoraphanin), indole glucosinolates (e.g., glucobrassicin), and aromatic glucosinolates (e.g., gluconasturtiin) (Figure 1) (1). Around 130 glucosinolate structures have been described to date (3), but only a subset can be found in the human diet. In a cohort of 2,121 German participants in the European Prospective Investigation into Cancer and Nutrition (EPIC study), glucobrassicin, sinigrin, glucoraphasatin (dehydroerucin), glucoraphanin, and glucoiberin were found to contribute most to total glucosinolate intake (4).
Glucosinolates and their breakdown derivatives (metabolites), especially isothiocyanates and indole-3-carbinol, exert a variety of biological activities that may be relevant to health promotion and disease prevention in humans (see the MIC articles on Indole-3-Carbinol and Isothiocyanates).
Metabolism and Bioavailability of Glucosinolates
Metabolism
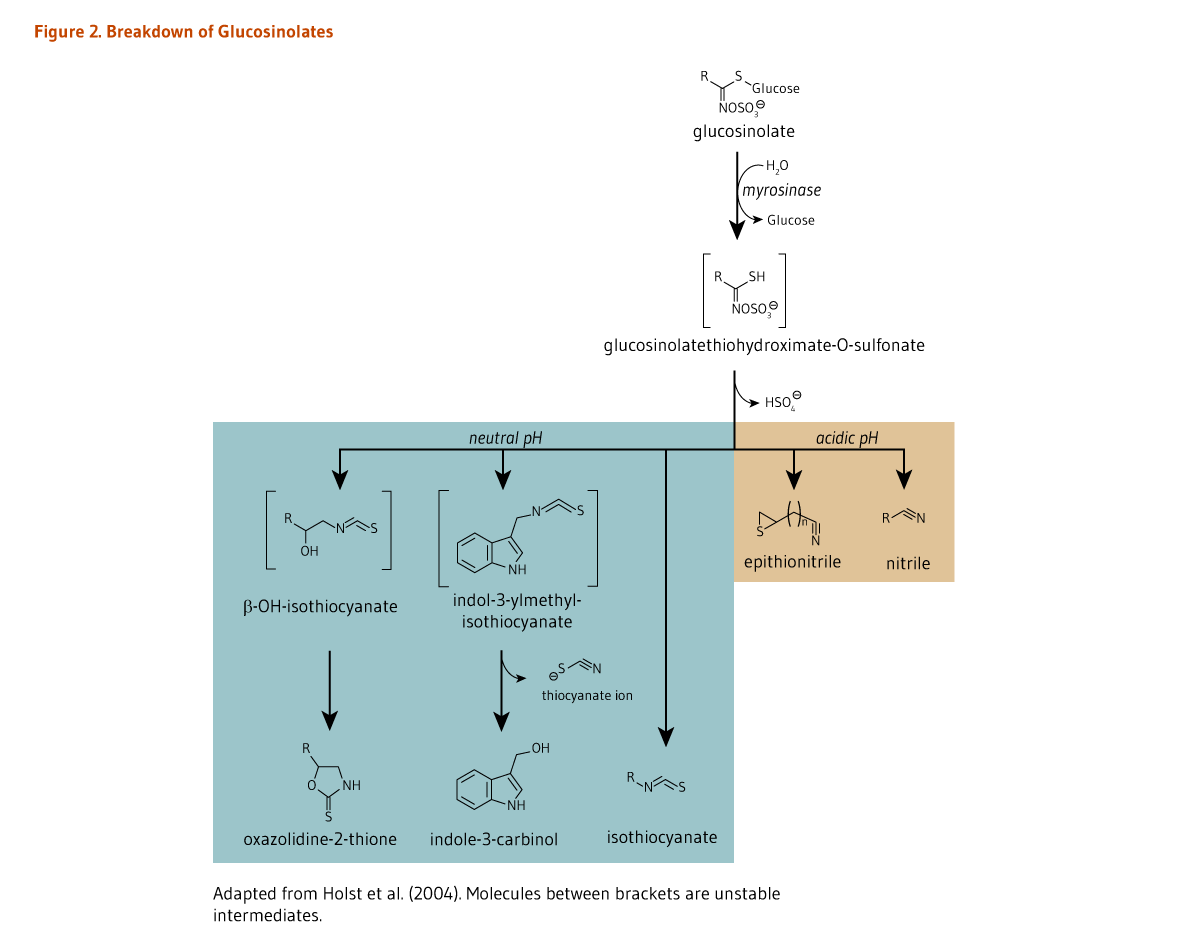
The hydrolysis of glucosinolates, which is catalyzed by a class of enzymes called myrosinases (β-thioglucosidases), leads to the formation of breakdown compounds, such as thiocyanates, isothiocyanates, indoles, oxazolidine-2-thiones (e.g., goitrin), epithionitrile, and nitrile (Figure 2). In intact plant cells, myrosinase is physically separated from glucosinolates. Yet, when plant cells are damaged, myrosinase is released and comes in contact with glucosinolates, catalyzing their conversion into highly reactive metabolites. In plants, thiocyanates, isothiocyanates, epithionitrile, and nitrile are defensive compounds against pathogens, insects, and herbivores (1). When raw cruciferous vegetables are chopped during the cooking process, glucosinolates are rapidly hydrolyzed by myrosinase, generating metabolites that are then absorbed in the proximal intestine. In contrast, boiling cruciferous vegetables before consumption inactivates myrosinase, thus preventing the breakdown of glucosinolates. A small fraction of intact glucosinolates may be absorbed in the small intestine, but a large proportion reaches the colon (5). Of note, boiling cruciferous vegetables has also been found to reduce their glucosinolate content to a much greater extent than steam cooking, microwaving, and stir-frying do (5). Nonetheless, when cruciferous vegetables are cooked, bacterial myrosinase-like activity in the colon is mainly responsible for glucosinolate degradation, generating a wide range of metabolites (5, 6).
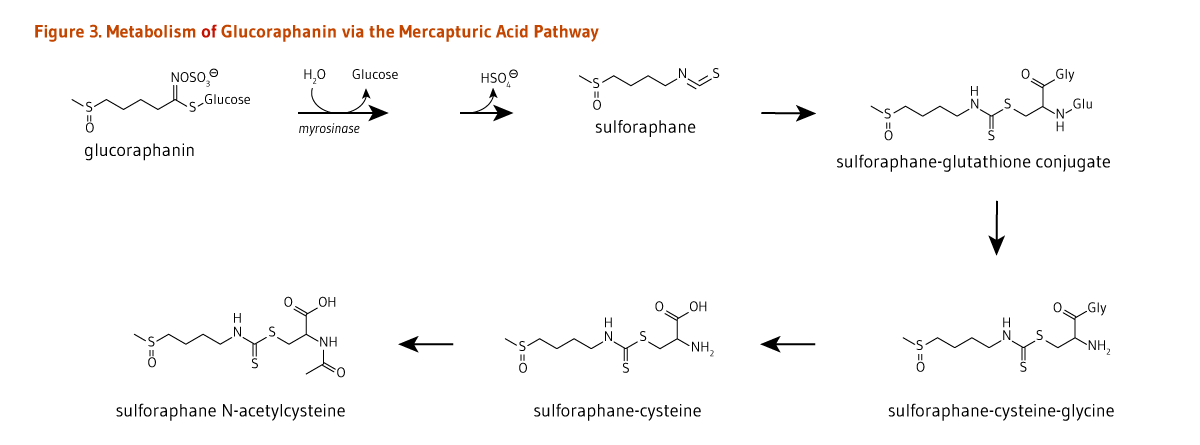
A neutral pH may favor the formation of isothiocyanates from glucosinolates (Figure 2). Once absorbed, isothiocyanates, such as glucoraphanin-derived sulforaphane, are conjugated to glutathione in the liver, and then sequentially metabolized in the mercapturic acid pathway (Figure 3). Sulforaphane metabolites — sulforaphane-glutathione, sulforaphane-cysteine-glycine, sulforaphane-cysteine, and sulforaphane N-acetylcysteine (Figure 3) — collectively known as dithiocarbamates, are ultimately excreted in the urine (5).
Bioavailability
The composition and content of glucosinolates in cruciferous vegetables are relatively stable, yet depend on the genus and species and can vary with plant growing and post-harvest storage conditions and culinary processing (7, 8). Since most cruciferous vegetables are cooked prior to eating, bacterial myrosinase-like activity in the gut rather than plant myrosinase is responsible for the initial step in glucosinolate degradation (Figure 2). In a feeding study involving 45 healthy subjects, the mean conversion rate of glucosinolates (of which 85% was glucoraphanin) to dithiocarbamates over a 24-hour period was estimated to be around 12% with wide variations among participants (range, 1.1 to 40.7%) (7). In contrast, 70%-75% of ingested isothiocyanates were found to be metabolized to dithiocarbamates. Therefore, following the ingestion of cooked cruciferous vegetables, the conversion of glucosinolates into isothiocyanates by gut bacteria appears to be a limiting step in the generation of dithiocarbamates (7). However, differences in individuals’ capacity to metabolize glucosinolates have not been linked to differences in gut microbiota composition (9).
Disease Prevention
Like most other vegetables, cruciferous vegetables are good sources of a variety of nutrients and phytochemicals that synergistically contribute to health promotion (see Bioactive compounds in cruciferous vegetables) (10). One challenge in studying the relationships between cruciferous vegetable intake and disease risk in humans is dissociating the benefits of whole diets that are generally rich in vegetables from those that are specifically rich in cruciferous vegetables (11). One characteristic that sets cruciferous vegetables apart from other vegetables is their high glucosinolate content (see Introduction). Glucosinolate hydrolysis products may play important roles in disease prevention by triggering antioxidant and anti-inflammatory response and contributing to the maintenance of cell homeostasis (see the MIC articles on Isothiocyanates and Indole-3-Carbinol).
Genetic influences
Once absorbed, glucosinolate-derived isothiocyanates (like sulforaphane) are promptly conjugated to glutathione by a class of phase II detoxification enzymes known as glutathione S-transferases (GSTs) (Figure 3). This mechanism is meant to increase the solubility of isothiocyanates, thereby promoting a rapid excretion in the urine. Isothiocyanates are thought to play a prominent role in the potential anticancer and cardiovascular benefits associated with cruciferous vegetable consumption (12, 13). Genetic variations in the sequence of genes coding for GSTs may affect the activity of these enzymes. Such variations have been identified in humans. Specifically, null variants of the GSTM1 and GSTT1 alleles contain large deletions, and individuals who inherit two copies of the GSTM1-null or GSTT1-null alleles cannot produce the corresponding GST enzymes (14). It has been proposed that a reduced GST activity in these individuals would slow the rate of excretion of isothiocyanates, thereby increasing tissue exposure to isothiocyanates after cruciferous vegetable consumption (15). However, human interventional studies with watercress report there is no difference in the isothiocyanate excretion rate between positive (+/+) and null (-/-) genotypes (16). Similar studies with broccoli have shown that GSTM1-/- individuals excreted a greater proportion of ingested sulforaphane via mercapturic acid metabolism than GSTM1+/+ individuals (17, 18). In addition, GSTs are involved in "detoxifying" potentially harmful substances like carcinogens, suggesting that individuals with reduced GST activity might also be more susceptible to cancer (19-21). Finally, induction of the expression and activity of GSTs and other phase II detoxification/antioxidant enzymes by isothiocyanates is an important defense mechanism against oxidative stress and damage associated with the development of diseases like cancer and cardiovascular disease (22). The ability of sulforaphane (glucoraphanin-derived isothiocyanate) to reduce oxidative stress in different settings is linked to activation of the nuclear factor E2-related factor 2 (Nrf2)-dependent pathway. Yet, whether potential protection conferred by isothiocyanates via the Nrf2-dependent pathway is diminished in individuals carrying GST-/- variants is currently unknown.
Some, but not all, observational studies have found that GST genotypes could influence the associations between isothiocyanate intake from cruciferous vegetables and risk of disease (23).
Cardiovascular disease
High intakes of fruit and vegetables have been consistently associated with a reduced risk of cardiovascular disease (CVD) (24, 25). Yet, few observational studies have specifically examined the potential benefits of cruciferous vegetable consumption. In the Shanghai Women’s Health Study (mean follow-up, 10.2 years) and the Shanghai Men’s Health Study (mean follow-up, 4.6 years), which included a total of 134,796 Chinese adults, participants in the highest versus lowest quintile of cruciferous vegetable intakes had a 22% reduced risk of all cause-mortality and a 31% reduced risk of CVD-related mortality (26). In contrast, a pooled analysis of two large US prospective cohort studies, the Nurses’ Health Study (70,870 women) and the Health Professionals’ Follow-Up Study (38,918 men), found no significant association between cruciferous vegetable intake and combined risk of myocardial infarction (MI) and ischemic stroke (27). A case-control study conducted in 2,042 subjects (ages, <75 years) who survived a first acute myocardial infarction (MI), and matched healthy controls with no CVD history found that the individuals in the highest versus lowest tertile of cruciferous vegetable intakes (6 times/week versus <1 time/week) had 27% lower odds of MI (28). However, further analyses showed that the association between cruciferous vegetable intake and MI events was significant in individuals with two functional GSTT1 alleles but not in carriers of two alleles of the GSTT1 null variant (-/-) (28).
Analysis of data from two 12-week randomized controlled trials in 130 participants with mild or moderate CVD risk found that the consumption of 400 g/week of high-glucosinolate broccoli (containing 3 to 6 times more glucoraphanin and glucoiberin than standard broccoli) resulted in a significant reduction in low-density lipoprotein (LDL)-cholesterol concentration in plasma compared with standard broccoli (29). Whether the effect of glucosinolates on cholesterol metabolism might be beneficial in the prevention of CVD needs further investigation.
Cancer
A recent intervention study demonstrated that cruciferous vegetables could increase the detoxification of carcinogens and other xenobiotics in humans. In this 12-week randomized controlled trial in 391 healthy Chinese adults exposed to high levels of air pollution, daily consumption of a broccoli sprout-rich beverage (providing 600 µmol/day of glucoraphanin and 40 µmol/day of sulforaphane) significantly increased the urinary excretion of a known carcinogen, benzene, and a toxicant, acrolein, compared to placebo (20). The biological activities of glucosinolate derivatives, isothiocyanates and indole-3-carbinol, which include modulation of xenobiotic metabolism, but also antioxidant and anti-inflammatory properties, induction of cell cycle arrest and apoptosis, and inhibition of angiogenesis, likely contribute to the potential benefits of cruciferous vegetables in the prevention of cancer (see the MIC articles on Isothiocyanates and Indole-3-Carbinol) (23).
Evidence from observational studies
Numerous observational studies have examined the relationship between cruciferous vegetable intake and cancer risk. Results from recent published meta-analyses of observational studies are reported in Table 1 (adapted from 23).
| Type of Cancer | Type of Observational Studies | Relative Risk [RR] or Odds Ratio [OR] (95% Confidence Interval) | Relative Risk [RR] in Subgroup Analyses (e.g., by food group or study type) | References |
|---|---|---|---|---|
| Bladder cancer | 5 prospective cohort and 5 case-control studies | RR: 0.80 (0.69-0.92) | RR: 0.78 (0.67-0.89) with case-control studies only RR: 0.86 (0.61-1.11) with cohort studies only |
Liu et al. (2013) (30) |
| 12 prospective cohort and case-control studies | RR: 0.84 (0.77-0.91) | Yao et al. (2014) (31) | ||
| 7 prospective cohort and case-control studies | RR: 0.85 (0.69-1.06) | Vieira et al. (2015) (32) | ||
| 8 prospective cohort studies | RR: 0.97 (0.93-1.01) | Xu et al. (2015) (33) | ||
| Breast cancer | 11 case-control studies | RR: 0.85 (0.77-0.94) | Liu et al. (2013) (34) | |
| Colorectal cancer | 24 case-control and 11 prospective cohort studies | RR: 0.82 (0.75-0.90) | RR: 0.76 (0.60-0.97) for studies reporting on cabbage intake RR: 0.82 (0.65-1.02) for studies reporting on broccoli intake |
Wu et al. (2013) (35) |
| 11 prospective cohort and 18 case-control studies | OR: 0.92 (0.83-1.01) | OR: 0.84 (0.72-0.98) for colon cancer OR: 0.99 (0.67-1.46) for rectal cancer OR: 1.09 (0.90-1.33) for colonic adenoma OR: 0.80 (0.65-0.99) for studies reporting on broccoli intake OR: 0.95 (0.80-1.14) for studies reporting on cabbage intake OR: 1.0 (0.75-1.34) for studies reporting on Brussels sprouts |
Tse et al. (2014) (36) | |
| Endometrial cancer | 1 prospective cohort study and 16 case-control studies | OR: 0.79 (0.69-0.90) per 100 g/day | Bandera et al. (2007) (37) | |
| Gastric cancer | 6 prospective cohort and 16 case-control studies | RR: 0.81 (0.75-0.88) | RR: 0.78 (0.71-0.86) for case-control studies RR: 0.89 (0.77-1.02) for cohort studies RR: 0.68 (0.58-0.80) for studies reporting on cabbage intake |
Wu et al. (2013) (38) |
| Lung cancer | 6 prospective cohort and 13 case-control studies | RR: 0.77 (0.68-0.88) for case-control studies RR: 0.83 (0.62-1.08) for cohort studies |
Lam et al. (2009) (39) | |
| 5 prospective cohort and 6 case-control studies | RR: 0.75 (0.63-0.89) | Wu et al. (2013) (40) | ||
| Ovarian cancer | 5 prospective cohort and 6 case-control studies | RR: 0.90 (0.82-0.98) | RR: 0.84 (0.75-0.94) for case-control studies RR: 1.0 (0.85-1.11) for cohort studies |
Han et al. (2014) (41) |
| 4 prospective cohort and 4 case-control studies | RR: 0.89 (0.81-0.99) | Hu et al. (2015) (42) | ||
| Pancreatic cancer | 4 prospective cohort and 5 case-control studies | OR: 0.79 (0.64-0.91) | OR: 0.72 (0.55-0.89) for case-control studies OR: 0.87 (0.67-1.06) for cohort studies OR: 0.78 (0.55-1.01) for high-quality studies OR: 0.80 (0.66-0.94) for low-quality studies |
Li et al. (2015) (43) |
| Prostate cancer | 7 prospective cohort and 6 case-control studies | RR: 0.90 (0.85-0.96) | RR: 0.79 (0.69-0.89) for case-control studies RR: 0.95 (0.89-1.02) for cohort studies |
Liu et al. (2012) (44) |
| Renal cell carcinoma | 6 prospective cohort and 6 case-control studies | RR: 0.81 (0.72-0.91) | RR: 0.89 (0.82-0.98) for high-quality studies RR: 0.72 (0.64-0.81) for case-control studies RR: 0.92 (0.84-1.00) for cohort studies |
Zhao et al. (2013) (45) |
| 3 prospective cohort and 7 case-control studies | RR: 0.73 (0.63-0.83) | RR: 0.69 (0.60-0.78) for case-control studies RR: 0.96 (0.71-1.21) for cohort studies |
Liu et al. (2013) (46) |
Most meta-analyses found inverse associations between cruciferous vegetable intake and risk of bladder, breast, colorectal, endometrial, gastric, lung, ovarian, pancreatic, prostate, and renal cancer. Subgroup analyses showed that inverse associations remained significant in pooled analyses of case-control studies but not in pooled analyses of prospective cohort studies (see Table 1). Retrospective case-control studies are susceptible to bias in the selection of participants (cases and controls) and prone to dietary recall bias compared to prospective cohort studies, which collect dietary information from participants before they are diagnosed with cancer (47). The method of cooking cruciferous vegetables, which strongly affects the bioavailability and potential anticancer benefits of isothiocyanates (see Metabolism and Bioavailability of Glucosinolates) may be a source of bias and explain variation in the results of the studies (heterogeneity among studies). The lack of information regarding cooking methods prevented data adjustment to reduce bias.
In the past decades, some observational studies have examined the effect of individuals’ genetic variations on the relationship between cruciferous vegetable intake and the risk of different cancer types. For example, a pooled analysis of two prospective cohort studies and six case-control studies found an inverse association between cruciferous vegetable consumption and risk of colorectal neoplasm in carriers of the GSTT1 null variant but not in individuals with the GSTM1 null variant or those with both the GSTT1 and GSTM1 null variants (-/-) (36). The results of a pooled analysis of five case-control studies also suggested a stronger association between cruciferous vegetable intake and lung cancer in carriers of both the GSTT1-/- and GSTM1-/- variants compared to carriers of wild-type alleles (+/+); however, it was not reported whether results from these two groups of individuals were significantly different (39). There is also a significant body of evidence suggesting that GSTM1+/+ individuals gain greater cancer protection from consumption total cruciferous vegetables or broccoli compared to GSTM1-/- variant carriers (25, 48, 49). Current evidence is scarce, and adequately powered, well-designed studies are required to assess and explain potential interactions between cruciferous vegetable intake and GST genotypes.
A few observational studies have looked at whether cruciferous vegetable intake could be associated with reduced risks of disease progression and mortality. The highest versus lowest intake of cruciferous vegetables (assessed before diagnosis) was associated with a better survival rate over 72 months after diagnosis in 547 women with lung cancer (50). A prospective study in 29,361 men who underwent a prostate-specific antigen (PSA) test found that intake of cruciferous vegetables was inversely associated with risk of metastatic prostate cancer — cancer that has spread beyond the prostate (i.e., late-stage prostate cancer) — during a mean follow-up of 4.2 years (51). Another prospective study in 1,560 men diagnosed with non-metastatic prostate cancer reported that higher post-diagnosis intake of cruciferous vegetables was associated with a 59% lower risk of prostate cancer progression during a two-year period after completion of the dietary assessment (52). In contrast, cruciferous vegetable consumption in a cohort of 11,390 women with stage I-III invasive breast cancer (from four US and Chinese prospective studies), assessed about two years after diagnosis, was not found to be associated with risk of cancer recurrence or total mortality (53).
Nutrient Interactions
Iodine and thyroid function
Very high intakes of cruciferous vegetables, such as cabbage and turnips, have been found to cause hypothyroidism (insufficient production of thyroid hormones) in animals (54). Two mechanisms can potentially explain this effect. The hydrolysis of progoitrin, found in cruciferous vegetables (see Figure 1), may yield a compound known as goitrin, which may interfere with thyroid hormone synthesis. The hydrolysis of another class of glucosinolates, known as indole glucosinolates, results in the release of thiocyanate ions (see Figure 2) that can compete with iodine for uptake by the thyroid gland (55). However, increased exposure to thiocyanate ions from cruciferous vegetable consumption or, more commonly, from cigarette smoking, does not appear to increase the risk of hypothyroidism unless accompanied by iodine deficiency. One study in humans found that the consumption of 150 g/day (5 oz/day) of cooked Brussels sprouts for four weeks had no adverse effects on thyroid function (56). Similarly, consumption of high amounts of cruciferous vegetables has been associated with increased thyroid cancer risk only in iodine-deficient areas (57).
Intake Recommendations
The 2015-2020 Dietary Guidelines for Americans recommend eating a variety of vegetables daily (2½ cup-equivalents/day for a 2,000 calorie diet) from all of the five vegetable subgroups (dark green, red and orange, legumes, starchy, and other; see 58). No separate recommendations have been established for cruciferous vegetables, yet the 2015-2020 Dietary Guidelines for Americans recommend that adults consume 1½-2½ cup-equivalents of dark-green vegetables (which include cruciferous vegetables) per week (58).
Bioactive compounds in cruciferous vegetables
Cruciferous vegetables are important sources of some vitamins and minerals, fiber, and various phytochemicals other than glucosinolates (Table 2). Many of these compounds likely contribute to the potential health-promoting benefits of cruciferous vegetables.
| Vitamins | Minerals | Phytochemicals |
|---|---|---|
| Folate | Potassium | Carotenoids |
| Vitamin C | Selenium | Chlorophyll |
| Vitamin K | Calcium | Fiber |
| Flavonoids | ||
| Indole-3-Carbinol | ||
| Isothiocyanates | ||
| Lignans | ||
| Phytosterols | ||
| Sulfur bioactives (other than glucosinolates) (59) |
Authors and Reviewers
Originally written in 2005 by:
Jane Higdon, Ph.D.
Linus Pauling Institute
Oregon State University
Updated in December 2008 by:
Victoria J. Drake, Ph.D.
Linus Pauling Institute
Oregon State University
Updated in December 2016 by:
Barbara Delage, Ph.D.
Linus Pauling Institute
Oregon State University
Reviewed in April 2017 by:
Maria Traka, Ph.D.
Senior Research Scientist
Chair of the Athena SWAN SAT
Food and Health Programme
Institute of Food Research
Norwich, United Kingdom
Copyright 2005-2026 Linus Pauling Institute
Figure Alternative Text
Figure 1. Chemical structures of some glucosinolates
The figure shows chemical structures of some glucosinolates and examples from three subclasses: aliphatic, aromatic, and indole glucosinolates. At the top left, the general glucosinolate structure is shown. Five different aliphatic glucosinolates are shown: sinigrin (2-propenylglucosinolate), glucoiberin (3-methylsulfinylpropylglucosinolate), glucoraphasatin (dehydroerucin; 4-methylthio-3-butenylglucosinolate), glucoraphanin (4-methylsulfinylbutylglucosinolate), and progoitrin (2-hydroxy-3-butenylglucosinolate). The figure legend states that aliphatic glucosinolates are derived from alanine, leucine, isoleucine, valine, and methionine.
At the bottom, two aromatic glucosinolates are shown: glucotropaeolin (benzylglucosinolate) and gluconasturtiin (2-phenylethylglucosinolate). The figure legend states that aromatic glucosinolates are derived from phenylalanine or tyrosine.
Also shown is an indole glucosinolate, glucobrassicin (3-indolylmethylglucosinolate), derived from tryptophan.
Figure 2. Breakdown of glucosinolates
The figure shows the biochemical breakdown of glucosinolates, including the chemical structures of the metabolites. At the top, glucosinolate is hydrolyzed by myrosinase, releasing glucose and forming the unstable intermediate, glucosinolatethiohydroximate-O-sulfonate. This intermediate can enter two pathways depending on pH. Under neutral pH (shown in a blue box), the intermediate is metabolized to a general isothiocyanate or to indole-3-carbinol or oxazolidine-2-thione via the unstable intermediates, indol-3-ylmethyl-isothiocyanate or β-OH-isothiocyanate, respectively. Under acidic pH (shown in a brown box), the products are epithionitrile and a general nitrile.
Figure 3. Metabolism of glucoraphanin via the mercapturic acid pathway
The figure shows the names and chemical structures of compounds and the enzymes involved. Glucoraphanin is hydrolyzed by myrosinase, releasing glucose and a hydrogen sulfate ion and forming sulforaphane. Sulforaphane undergoes conjugation by glutathione S-transferase, forming a sulforaphane-glutathione conjugate. This conjugate is metabolized by γ-glutamyltranspeptidase into sulforaphane-cysteine-glycine, then by cysteinyl-glycinase to form sulforaphane-cysteine, and finally by N-acetyltransferase to form sulforaphane N-acetylcysteine.
References
- Ishida M, Hara M, Fukino N, Kakizaki T, Morimitsu Y. Glucosinolate metabolism, functionality and breeding for the improvement of Brassicaceae vegetables. Breed Sci. 2014;64(1):48-59. (PubMed)
- International Agency for Research on Cancer. Cruciferous vegetables. Cruciferous vegetables, isothiocyanates and indoles. Lyon, France: IARC; 2004:1-12.
- Agerbirk N, Olsen CE. Glucosinolate structures in evolution. Phytochemistry. 2012;77:16-45. (PubMed)
- Steinbrecher A, Linseisen J. Dietary intake of individual glucosinolates in participants of the EPIC-Heidelberg cohort study. Ann Nutr Metab. 2009;54(2):87-96. (PubMed)
- Barba FJ, Nikmaram N, Roohinejad S, Khelfa A, Zhu Z, Koubaa M. Bioavailability of glucosinolates and their breakdown products: impact of processing. Front Nutr. 2016;3:24. (PubMed)
- Luang-In V, Albaser AA, Nueno-Palop C, Bennett MH, Narbad A, Rossiter JT. Glucosinolate and desulfo-glucosinolate metabolism by a selection of human gut bacteria. Curr Microbiol. 2016;73(3):442-451. (PubMed)
- Fahey JW, Wehage SL, Holtzclaw WD, et al. Protection of humans by plant glucosinolates: efficiency of conversion of glucosinolates to isothiocyanates by the gastrointestinal microflora. Cancer Prev Res (Phila). 2012;5(4):603-611. (PubMed)
- Verkerk R, Schreiner M, Krumbein A, et al. Glucosinolates in Brassica vegetables: the influence of the food supply chain on intake, bioavailability and human health. Mol Nutr Food Res. 2009;53 Suppl 2:S219. (PubMed)
- Li F, Hullar MA, Beresford SA, Lampe JW. Variation of glucoraphanin metabolism in vivo and ex vivo by human gut bacteria. Br J Nutr. 2011;106(3):408-416. (PubMed)
- Liu RH. Potential synergy of phytochemicals in cancer prevention: mechanism of action. J Nutr. 2004;134(12 Suppl):3479S-3485S. (PubMed)
- McNaughton SA, Marks GC. Development of a food composition database for the estimation of dietary intakes of glucosinolates, the biologically active constituents of cruciferous vegetables. Br J Nutr. 2003;90(3):687-697. (PubMed)
- Bai Y, Wang X, Zhao S, Ma C, Cui J, Zheng Y. Sulforaphane protects against cardiovascular disease via Nrf2 activation. Oxid Med Cell Longev. 2015;2015:407580. (PubMed)
- Higdon JV, Delage B, Williams DE, Dashwood RH. Cruciferous vegetables and human cancer risk: epidemiologic evidence and mechanistic basis. Pharmacol Res. 2007;55(3):224-236. (PubMed)
- Coles BF, Kadlubar FF. Detoxification of electrophilic compounds by glutathione S-transferase catalysis: determinants of individual response to chemical carcinogens and chemotherapeutic drugs? Biofactors. 2003;17(1-4):115-130. (PubMed)
- Seow A, Shi CY, Chung FL, et al. Urinary total isothiocyanate (ITC) in a population-based sample of middle-aged and older Chinese in Singapore: relationship with dietary total ITC and glutathione S-transferase M1/T1/P1 genotypes. Cancer Epidemiol Biomarkers Prev. 1998;7(9):775-781. (PubMed)
- Dyba M, Wang A, Noone AM, et al. Metabolism of isothiocyanates in individuals with positive and null GSTT1 and M1 genotypes after drinking watercress juice. Clin Nutr. 2010;29(6):813-818. (PubMed)
- Gasper AV, Al-Janobi A, Smith JA, et al. Glutathione S-transferase M1 polymorphism and metabolism of sulforaphane from standard and high-glucosinolate broccoli. Am J Clin Nutr. 2005;82(6):1283-1291. (PubMed)
- Steck SE, Gammon MD, Hebert JR, Wall DE, Zeisel SH. GSTM1, GSTT1, GSTP1, and GSTA1 polymorphisms and urinary isothiocyanate metabolites following broccoli consumption in humans. J Nutr. 2007;137(4):904-909. (PubMed)
- Economopoulos KP, Sergentanis TN. GSTM1, GSTT1, GSTP1, GSTA1 and colorectal cancer risk: a comprehensive meta-analysis. Eur J Cancer. 2010;46(9):1617-1631. (PubMed)
- Egner PA, Chen JG, Zarth AT, et al. Rapid and sustainable detoxication of airborne pollutants by broccoli sprout beverage: results of a randomized clinical trial in China. Cancer Prev Res (Phila). 2014;7(8):813-823. (PubMed)
- Sergentanis TN, Economopoulos KP. GSTT1 and GSTP1 polymorphisms and breast cancer risk: a meta-analysis. Breast Cancer Res Treat. 2010;121(1):195-202. (PubMed)
- Bryan HK, Olayanju A, Goldring CE, Park BK. The Nrf2 cell defence pathway: Keap1-dependent and -independent mechanisms of regulation. Biochem Pharmacol. 2013;85(6):705-717. (PubMed)
- Traka MH. Chapter nine - Health benefits of glucosinolates. Advances in Botanical Research. 2016;80:247-279.
- Nothlings U, Schulze MB, Weikert C, et al. Intake of vegetables, legumes, and fruit, and risk for all-cause, cardiovascular, and cancer mortality in a European diabetic population. J Nutr. 2008;138(4):775-781. (PubMed)
- Wang X, Ouyang Y, Liu J, et al. Fruit and vegetable consumption and mortality from all causes, cardiovascular disease, and cancer: systematic review and dose-response meta-analysis of prospective cohort studies. BMJ. 2014;349:g4490. (PubMed)
- Zhang X, Shu XO, Xiang YB, et al. Cruciferous vegetable consumption is associated with a reduced risk of total and cardiovascular disease mortality. Am J Clin Nutr. 2011;94(1):240-246. (PubMed)
- Joshipura KJ, Hung HC, Li TY, et al. Intakes of fruits, vegetables and carbohydrate and the risk of CVD. Public Health Nutr. 2009;12(1):115-121. (PubMed)
- Cornelis MC, El-Sohemy A, Campos H. GSTT1 genotype modifies the association between cruciferous vegetable intake and the risk of myocardial infarction. Am J Clin Nutr. 2007;86(3):752-758. (PubMed)
- Armah CN, Derdemezis C, Traka MH, et al. Diet rich in high glucoraphanin broccoli reduces plasma LDL cholesterol: Evidence from randomised controlled trials. Mol Nutr Food Res. 2015;59(5):918-926. (PubMed)
- Liu B, Mao Q, Lin Y, Zhou F, Xie L. The association of cruciferous vegetables intake and risk of bladder cancer: a meta-analysis. World J Urol. 2013;31(1):127-133. (PubMed)
- Yao B, Yan Y, Ye X, et al. Intake of fruit and vegetables and risk of bladder cancer: a dose-response meta-analysis of observational studies. Cancer Causes Control. 2014;25(12):1645-1658. (PubMed)
- Vieira AR, Vingeliene S, Chan DS, et al. Fruits, vegetables, and bladder cancer risk: a systematic review and meta-analysis. Cancer Med. 2015;4(1):136-146. (PubMed)
- Xu C, Zeng XT, Liu TZ, et al. Fruits and vegetables intake and risk of bladder cancer: a PRISMA-compliant systematic review and dose-response meta-analysis of prospective cohort studies. Medicine (Baltimore). 2015;94(17):e759. (PubMed)
- Liu X, Lv K. Cruciferous vegetables intake is inversely associated with risk of breast cancer: a meta-analysis. Breast. 2013;22(3):309-313. (PubMed)
- Wu QJ, Yang Y, Vogtmann E, et al. Cruciferous vegetables intake and the risk of colorectal cancer: a meta-analysis of observational studies. Ann Oncol. 2013;24(4):1079-1087. (PubMed)
- Tse G, Eslick GD. Cruciferous vegetables and risk of colorectal neoplasms: a systematic review and meta-analysis. Nutr Cancer. 2014;66(1):128-139. (PubMed)
- Bandera EV, Kushi LH, Moore DF, Gifkins DM, McCullough ML. Fruits and vegetables and endometrial cancer risk: a systematic literature review and meta-analysis. Nutr Cancer. 2007;58(1):6-21. (PubMed)
- Wu QJ, Yang Y, Wang J, Han LH, Xiang YB. Cruciferous vegetable consumption and gastric cancer risk: a meta-analysis of epidemiological studies. Cancer Sci. 2013;104(8):1067-1073. (PubMed)
- Lam TK, Gallicchio L, Lindsley K, et al. Cruciferous vegetable consumption and lung cancer risk: a systematic review. Cancer Epidemiol Biomarkers Prev. 2009;18(1):184-195. (PubMed)
- Wu QJ, Xie L, Zheng W, et al. Cruciferous vegetables consumption and the risk of female lung cancer: a prospective study and a meta-analysis. Ann Oncol. 2013;24(7):1918-1924. (PubMed)
- Han B, Li X, Yu T. Cruciferous vegetables consumption and the risk of ovarian cancer: a meta-analysis of observational studies. Diagn Pathol. 2014;9:7. (PubMed)
- Hu J, Hu Y, Hu Y, Zheng S. Intake of cruciferous vegetables is associated with reduced risk of ovarian cancer: a meta-analysis. Asia Pac J Clin Nutr. 2015;24(1):101-109. (PubMed)
- Li LY, Luo Y, Lu MD, Xu XW, Lin HD, Zheng ZQ. Cruciferous vegetable consumption and the risk of pancreatic cancer: a meta-analysis. World J Surg Oncol. 2015;13:44. (PubMed)
- Liu B, Mao Q, Cao M, Xie L. Cruciferous vegetables intake and risk of prostate cancer: a meta-analysis. Int J Urol. 2012;19(2):134-141. (PubMed)
- Zhao J, Zhao L. Cruciferous vegetables intake is associated with lower risk of renal cell carcinoma: evidence from a meta-analysis of observational studies. PLoS One. 2013;8(10):e75732. (PubMed)
- Liu B, Mao Q, Wang X, et al. Cruciferous vegetables consumption and risk of renal cell carcinoma: a meta-analysis. Nutr Cancer. 2013;65(5):668-676. (PubMed)
- Song JW, Chung KC. Observational studies: cohort and case-control studies. Plast Reconstr Surg. 2010;126(6):2234-2242. (PubMed)
- Joseph MA, Moysich KB, Freudenheim JL, et al. Cruciferous vegetables, genetic polymorphisms in glutathione S-transferases M1 and T1, and prostate cancer risk. Nutr Cancer. 2004;50(2):206-213. (PubMed)
- Spitz MR, Duphorne CM, Detry MA, et al. Dietary intake of isothiocyanates: evidence of a joint effect with glutathione S-transferase polymorphisms in lung cancer risk. Cancer Epidemiol Biomarkers Prev. 2000;9(10):1017-1020. (PubMed)
- Wu QJ, Yang G, Zheng W, et al. Pre-diagnostic cruciferous vegetables intake and lung cancer survival among Chinese women. Sci Rep. 2015;5:10306. (PubMed)
- Kirsh VA, Peters U, Mayne ST, et al. Prospective study of fruit and vegetable intake and risk of prostate cancer. J Natl Cancer Inst. 2007;99(15):1200-1209. (PubMed)
- Richman EL, Carroll PR, Chan JM. Vegetable and fruit intake after diagnosis and risk of prostate cancer progression. Int J Cancer. 2012;131(1):201-210. (PubMed)
- Nechuta S, Caan BJ, Chen WY, et al. Postdiagnosis cruciferous vegetable consumption and breast cancer outcomes: a report from the After Breast Cancer Pooling Project. Cancer Epidemiol Biomarkers Prev. 2013;22(8):1451-1456. (PubMed)
- Fenwick GR, Heaney RK, Mullin WJ. Glucosinolates and their breakdown products in food and food plants. Crit Rev Food Sci Nutr. 1983;18(2):123-201. (PubMed)
- Felker P, Bunch R, Leung AM. Concentrations of thiocyanate and goitrin in human plasma, their precursor concentrations in brassica vegetables, and associated potential risk for hypothyroidism. Nutr Rev. 2016;74(4):248-258. (PubMed)
- McMillan M, Spinks EA, Fenwick GR. Preliminary observations on the effect of dietary brussels sprouts on thyroid function. Hum Toxicol. 1986;5(1):15-19. (PubMed)
- Cho YA, Kim J. Dietary factors affecting thyroid cancer risk: a meta-analysis. Nutr Cancer. 2015;67(5):811-817. (PubMed)
- US Department of Health and Human Services and US Department of Agriculture. 2015-2020 Dietary Guidelines for Americans. 8th ed.; 2015.
- Traka MH, Saha S, Huseby S, et al. Genetic regulation of glucoraphanin accumulation in Beneforte broccoli. New Phytol. 2013;198(4):1085-1095. (PubMed)
Disclaimer
The Linus Pauling Institute's Micronutrient Information Center provides scientific information on the health aspects of dietary factors and supplements, food, and beverages for the general public. The information is made available with the understanding that the author and publisher are not providing medical, psychological, or nutritional counseling services on this site. The information should not be used in place of a consultation with a competent health care or nutrition professional.
The information on dietary factors and supplements, food, and beverages contained on this website does not cover all possible uses, actions, precautions, side effects, and interactions. It is not intended as nutritional or medical advice for individual problems. Liability for individual actions or omissions based upon the contents of this site is expressly disclaimed.
You may not copy, modify, distribute, display, transmit, perform, publish or sell any of the copyrightable material on this website. You may hyperlink to this website but must include the following statement:
"This link leads to a website provided by the Linus Pauling Institute at Oregon State University. [Your name] is not affiliated or endorsed by the Linus Pauling Institute or Oregon State University."