Contenido
El selenio es un elemento traza que es esencial en pequeñas cantidades, pero como todos los elementos esenciales, el selenio puede ser tóxico en niveles elevados. A diferencia de las plantas, la mayoría de los animales — incluyendo a los seres humanos — requieren selenio para el funcionamiento apropiado de un cierto número de enzimas dependientes de selenio conocidas como selenoproteínas. Durante la síntesis de proteínas (traducción) el aminoácido selenocisteína es incorporado en proteínas elongadas en ubicaciones muy específicas en la secuencia de aminoácidos con el objetivo de formar selenoproteínas funcionales. Aunque plantas superiores no parecen necesitar selenio para su sobrevivencia, estas pueden incorporar lo de manera no específica en moléculas que contienen sulfuro cuando el mineral está presente en el suelo (1). Debe notarse que, en los animales, el aminoácido selenometionina puede ser incorporado de manera no especifica en las proteínas en lugar de metionina (2). Sin embargo, solo las proteínas que contienen selenocisteína son consideradas como selenoproteínas (Figura 1).

[Figura 1 - Clic para Agrandar]
Veinticinco genes que codifican para las selenoproteínas han sido identificados en los humanos (3). La inserción de selenocisteína en las selenoproteínas durante la traducción es dirigida por la presencia de una secuencia de inserción de selenocisteína (SECIS) dentro de los ARNm de la selenoproteína. Brevemente, el reconocimiento de SECIS por la maquinaria de traducción resulta en el reclutamiento de factores de traducción específicos que decodifican codones UGA en marco al insertar selenocisteína dentro de selenoproteínas elongadas (4).
La investigación esta gradualmente descubriendo las funciones metabólicas de todas las selenoproteínas humanas, incluyendo variantes de empalme (3). Algunas de las selenoproteínas con una función identificada incluyen:
Se han identificado cinco enzimas glutatión peroxidasas (GPx1-4 y GPx6) que contienen selenio: la GPx1 (GPx citosólica), la GPx2 (GPx especifica de las células epiteliales expresada en el recubrimiento intestinal y pulmones) la GPx3 (altamente expresada en las glándulas tiroideas y rinones), la GPx4 (GPx fosfolípido-hidroperóxido; PHGPx) y la Gpx6 (expresada en el epitelio olfativo) (4). Las isoenzimas GPx son todas enzimas antioxidantes que reducen a las especies reactivas de oxígeno (ERO) potencialmente dañinas, como el peróxido de hidrógeno y los hidroperóxidos lipídicos, en productos inocuos como el agua y los alcoholes al acoplar su reducción con la oxidación de glutatión (Figura 2). La espermatogénesis y la fertilidad masculina son altamente dependientes de GPx4 y de la selenoproteína P (SEPP1; véase abajo). En los testículos, la GPx4 reduce los hidroperóxidos fosfolípidos, por consiguiente, protegiendo espermatozoides inmaduros contra el estrés oxidativo. La GPx4 es también una proteína estructural principal de la capsula que incorpora la espiral mitocondrial del espermatozoide maduro involucrada en la motilidad del esperma. La SEPP1es esencial para el suministro de selenio a los testículos, y modelos animales carentes del gen SEPP1 son infértiles debido a la biodisponibilidad pobre de selenio en los tejidos, a la síntesis defectuosa de GPx4, y a la maduración deteriorada del esperma (5).

[Figura 2 - Clic para Agrandar]
En los mamíferos, tres isoenzimas tiorredoxina reductasas que contienen selenocisteína (TrxR) han sido identificadas en el sistema tiorredoxina: TrxR1 citosólica, TrxR3 mitocondrial, y tiorredoxina glutatión reductasa testículo-especifica TGR. Las TrxRs son enzimas homodiméricas, y cada monómero contiene dominios de unión FAD- y NADPH- y un sitio catalítico que contiene selenocisteína. Las TrxRs catalizan la reducción de un amplio rango de sustratos, incluyendo la tiorredoxina y la proteína disulfuro isomerasa (PDI) (véase Figura 2 arriba). TrxRs también sirven como donadores de electrones para la regeneración de antioxidantes pequeños, reciclando posiblemente el ácido ascórbico (vitamina C), ácido α-lipoico, α-tocoferol (vitamina E) y la coenzima Q10 de sus formas oxidadas (6). El mantenimiento de la tiorredoxina en una forma reducida por las TrxRs es importante para la regulación del crecimiento celular y la supervivencia. La proteína tiorredoxina, junto con TrxR1 (o TrxR3), NADPH, y FAD, constituyen el sistema antioxidante tiorredoxina involucrado en la reducción de enzimas antioxidantes (p. ej., peroxirredoxinas, metionina sulfóxido reductasas y ribonucleótido reductasa) y de muchas proteínas de señalización sensibles a la oxidación/reducción (redox) (7). La TrxR1 es una de las selenoproteínas más investigadas y es considerada una de las principales enzimas antioxidantes y reguladoras del proceso redox en células mamíferas.
La glándula tiroides libera cantidades muy pequeñas de la forma biológicamente activa de la hormona tiroidea (triyodotironina o T3) y grandes cantidades de la hormona tiroidea en su forma inactiva (precursor de la T3: tiroxina o T4) en la circulación. La mayoría de la T3 biológicamente activa en la circulación y dentro de las células es generada a través de la remoción de un átomo de yodo de la T4 en una reacción catalizada por enzimas iodotironina deiodinasas dependientes de selenio. Dos diferentes iodotironina deiodinasas dependientes de selenio (DIO tipo 1 y 2) pueden desiodar la T4, incrementando así la T3 circulante, mientras que una tercera iodotironina deiodinasa (DIO tipo 3) puede convertir ambas T3 y T4 a metabolitos inactivos (Figura 3) (8). Debe notarse que, la inactivación de genes que codifican para las DIO en modelos de roedores ha revelado un papel para la DIO tipo 1 en la homeostasis del yodo y la importancia de las DIO tipo 2 y 3 en la maduración de los sistemas auditorio y visual durante el desarrollo fetal (8). Así, la importancia del selenio en el desarrollo, crecimiento y metabolismo normal no está limitado a su papel en la regulación de la función de la glándula tiroides.
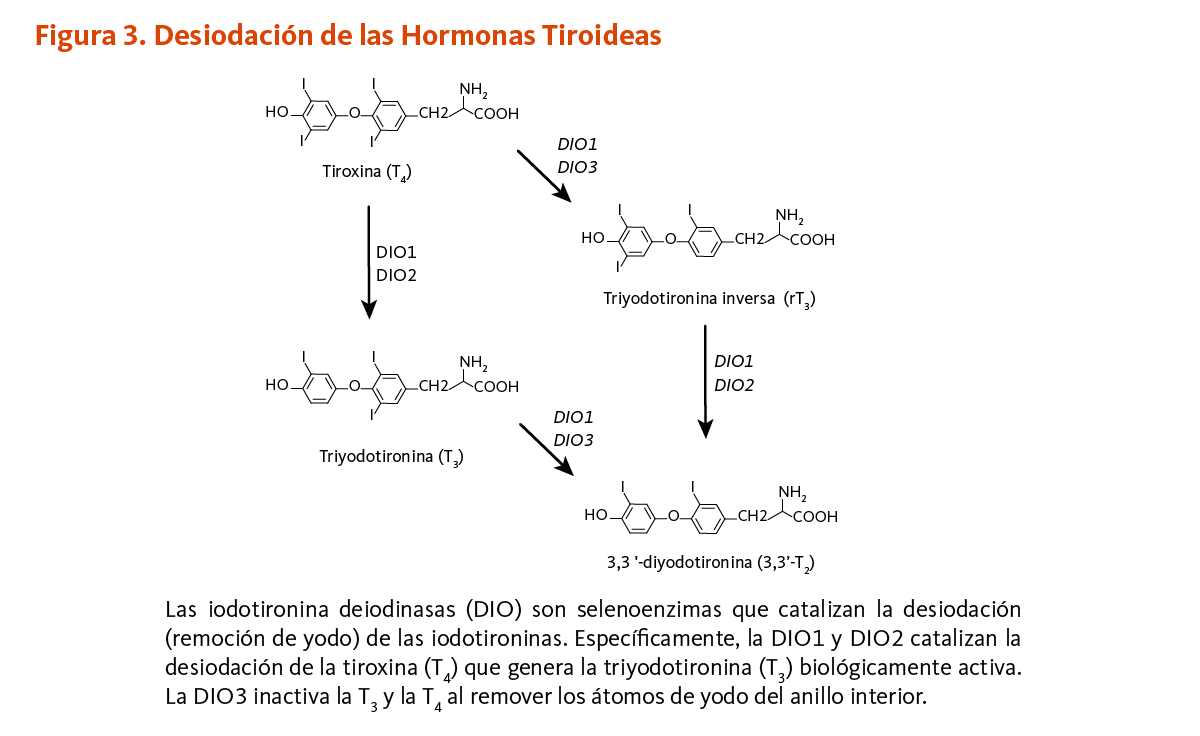
[Figura 3 - Clic para Agrandar]
La selenoproteína P (SEPP1) es predominantemente producida por el hígado, un sitio principal de almacenamiento para el selenio, y secretado en el plasma. La glicoproteína en toda su longitud contiene un dominio rico en selenio con nueve residuos de selenocisteína, como también un sitio catalítico similar a la tiorredoxina con un residuo de selenocisteína. La SEPP1 constituye la principal forma de transporte de selenio a los tejidos periféricos (9). SEPP1 también funciona como un antioxidante que protege a las células del daño oxidativo al permitir la completa actividad de las tiorredoxina reductasas y glutatión peroxidasas a través de un suministro adecuado de selenio a los tejidos extrahepáticos (véase Glutatión peroxidasas). SEPP1 parece ser especialmente crítica para la homeostasis del selenio en el cerebro y los testículos donde el receptor 2 de apolipoproteína E (apoER2) facilita la absorción de SEPP1. La megalina es otro receptor lipoproteico SEPP1-especifico que ayuda a limitar la perdida urinaria de selenio a través de la reabsorción de SEPP1 por los riñones (10). Además, la SEPP1 ha sido recientemente implicada en la regulación del metabolismo de la glucosa y la sensibilidad a la insulina (11).
La selenoproteína W (SEPW o SelW) existe en diferentes isoformas (homólogos) y es altamente conservada entre especies. En los humanos, la SEPW es expresada en numerosos tejidos, encontrando los niveles más altos en el musculo esquelético y corazón (12). SEPW contiene un residuo de selenocisteína y un residuo de cisteína que se une a una molécula de glutatión, sugiriendo un papel en la regulación del proceso redox (13). La expresión de SEPW esta correlacionada con el estatus del selenio y parece ser sensible a bajos suministros de selenio (14, 15). Se ha encontrado que la expresión de SEPW en el cerebro confiere protección contra la muerte de células neuronales inducida por el estrés oxidativo (16). La SEPW parece también ser un regulador negativo para las proteínas 14-3-3. Ciertamente, se encontró que la inhibición de 14-3-3 por la SEPW en las células del cáncer de seno incremento la proliferación celular y la supervivencia celular a través del incremento en la resistencia al estrés genotóxico (17). En las células del músculo esquelético, se demostró que SEPW redujo la unión de 14-3-3 a TAZ, permitiendo la translocación de TAZ al núcleo y la subsecuente activación de los genes de diferenciación celular del musculo (18). Finalmente, se encontró que la SEPW previene la degradación del receptor del factor de crecimiento epidérmico (EGFR) en células epiteliales del seno y próstata en cultivo. El EGFR es constitutivamente activado en muchos tumores, y evidencia de un papel para la SEPW en la activación y señalización del EGFR podría ayudar a arrojar luz sobre la relación entre el estatus del selenio y el riesgo de cáncer (19).
No existe un suministro libre del aminoácido selenocisteína en las células tal que la síntesis de selenocisteína tome lugar en un ARNt especializado durante la traducción del ARNm de la selenoproteína. La reacción es catalizada por la L-seril-ARNtSec selenio transferasa dependiente de piridoxal-5'-fosfato (PLP) y usa selenofosfato (monoselenio fosfato) como donante de selenio (Figura 4) (20). La selenofosfato sintetasa 2 es una selenoenzima que cataliza la síntesis dependiente de ATP del selenofosfato a partir del seleniuro de hidrógeno (Figura 4) (3).
![Figura 4. Síntesis de la Selenocisteína. La síntesis de selenocisteína (Sec) toma lugar en un ARNt especial inicialmente aminoacetilado (cargado) con el aminoácido serina (Ser). Ser-ARNt[Ser]Sec es entonces fosforilado para producir pSer-ARNt[Ser]Sec. La L-seril-ARNtSec selenio transferasa dependiente de piridoxal-5'-fosfato (PLP) cataliza la conversión de Ser-ARNt[Ser]Sec a Sec-ARNt[Ser]Sec usando selenofosfato como donante de selenio. El selenofosfato es sintetizado a partir del seleniuro de hidrógeno en una reacción catalizada por la selenoenzima selenofosfato sintetasa 2. La selenocisteína es incorporada en las proteinas elongadas durante la traducción. Se, selenio; Ser-ARNt, L-seril-ARNt; Sec-ARNt, L-selenocisteinilo-ARNt.](/sites/lpi.oregonstate.edu/files/selenio-figure-4-1200px.png)
[Figura 4 - Clic para Agrandar]
El sistema de reducción de la metionina sulfóxido está involucrado en la protección contra el estrés oxidativo y es especialmente crítico para la regeneración de proteínas dañadas por las especies reactivas de oxigeno (ERO). Ciertamente, las ERO pueden oxidar los residuos de metionina (sulfóxidos de metionina) dentro de las proteínas y potencialmente deteriorar sus actividades. En los humanos, dos familias estereospecíficas de las metionina sulfóxido reductasas (MsrA y MsrB) son codificadas por un único gen MSRA y tres genes MSRB (MSRB1-3). La MsrA cataliza la reducción de la forma-S de la metionina sulfóxido; la forma-R de la metionina sulfóxido es reducida por la MsrB, 2 y 3. Solo la MsrB1 ha sido caracterizada como una selenoproteína con un residuo de selenocisteína en su sitio catalítico. La MsrB1 parece estar involucrada en la regulación redox de ciertas proteínas. En los macrófagos, la reorganización de la actina del citoesqueleto necesaria para la quimiotaxis y fagocitosis requiere la reducción dependiente de MsrB1 de los residuos de metionina-R-sulfóxido dentro de la actina (21). Estudios que usaron la inactivación del gen MSR en ratones han mostrado que la reducción del sulfóxido de metionina está implicada en la regulación del ciclo de la metionina (revisado en 22). Se ha encontrado que tanto el sistema antioxidante de la tiorredoxina (Trx) como el de la glutarredoxina depediente de GSH (Grx) reducen las metionina sulfóxido reductasas in vitro y/o in vivo (véase Figura 2 arriba) (22).
La selenoproteína de 15 kDa (selenoproteína 15; SEP15) es altamente expresada en varios tejidos, incluyendo la próstata, riñones, hígado, y cerebro (23). Aunque su función no es aun conocida, se encontró que la SEP15 interactúa con la UDP-glucosa:glicoproteína glucosiltransferasa (UGGT) del retículo endoplásmico, una enzima involucrada en el control de calidad del plegamiento de las glicoproteínas (24, 25). Debido a que la SEP15 tiene un sitio catalítico como la tiorredoxina, se piensa que la SEP15 regula ya sea la actividad de la UGGT o el estado redox de los sustratos de UGGT (26). Se encontró que los ratones carentes de una SEP15 funcional desarrollaron cataratas nucleares (opacificación del cristalino) a una edad temprana sugiriendo que la SEP15 puede ser critica para el sistema de control de calidad del plegamiento de proteínas en el cristalino (27). La SEP15 puede también estar implicada en los mecanismos anticancerígenos (revisado en 28).
La selenoproteína mamífera (conocida como SEPS1, SelS, o selenoproteína de membrana que interactúa con VCP [VIMP]) es una proteína de membrana del retículo endoplasmático (RE). La SEPS1 está involucrada en la respuesta celular al estrés del RE (degradación asociada al RE; DARE) activada por la detección de proteínas mal plegadas. La SEP1 contribuye a la remoción y transferencia (retrotranslocalización) de las proteínas mal plegadas provenientes del lumen del RE al citosol donde las proteínas son marcadas con ubiquitina antes de ser degradadas. Se encontró que un polimorfismo o variación en la secuencia dentro de un elemento de respuesta al RE localizado en el promotor del SEPS1 resulta en la actividad disminuida del promotor del SEPS1 y de la expresión de genes (29). El polimorfismo correspondiente a la sustitución de una guanina (G) por una adenina (A) en el nucleótido -105 (-105G>A) ha sido asociado con niveles incrementados de citoquinas proinflamatorias en el plasma. Además, un estudio de caso y control recientemente reportó que el alelo A no era más prevalente en individuos afectados por la tiroiditis de Hashimoto (TH) — una enfermedad autoinmune mediada por células T resultando en la destrucción de las células tiroideas — que en los controles saludables (30). Otras asociaciones entre el polimorfismo del SEPS1 (incluyendo -105G>A) y la susceptibilidad a varias condiciones, como la preeclampsia, la enfermedad coronaria cardíaca, o los canceres gastrointestinales, sugieren fuertemente un papel para esta selenoproteína en la regulación de las respuestas inflamatorias e inmunes (31-34).
Otras selenoproteínas menos bien caracterizadas, que también son localizadas en el lumen del RE y/o la membrana, incluyen las selenoproteínas K, M, N y T (35).
La importancia del selenio para los sistemas biológicos, y específicamente para el balance redox celular (pro-oxidante/antioxidante), se deriva de su presencia como selenocisteína en el sitio catalítico de las selenoproteínas (véase Función). Otros minerales que son componentes críticos de las enzimas antioxidantes incluyen el cobre (como superóxido dismutasa), zinc (como superóxido dismutasa), y hierro (como catalasa). El selenio actúa en sinergia con las vitaminas antioxidantes, la vitamina C (ácido ascórbico) y la vitamina E (α-tocoferol), al regenerarlos a partir de sus formas oxidadas y promoviendo su protección antioxidante máxima (36-38).
Mientras que el yodo es un componente esencial de las hormonas tiroideas, las iodotironina deiodinasas que contienen selenio (DIO) son enzimas requeridas para la conversión de tiroxina (T4) a la hormona tiroidea biológicamente activa triyodotironina (T3) (véase Función). La actividad de la DIO1 puede también estar involucrada en la regulación de la homeostasis del yodo (39). Las selenoenzimas, glutatión peroxidasas, también desempeñan un papel crítico en la función tiroidea porque catalizan la degradación de peróxidos generados durante la síntesis de la hormona tiroidea (8). La epidemiologia de las deficiencias coexistentes de yodo y selenio en África central, pero no en China, ha sido ligada a la prevalencia de cretinismo mixedematoso, una forma severa de hipotiroidismo congénito acompañado por retraso mental y físico. La deficiencia de selenio puede ser solo uno de varios factores indeterminados que podrían exacerbar los efectos perjudiciales de la deficiencia de yodo (40). Curiosamente, se encontró que la deficiencia de selenio en roedores tiene poco impacto en las actividades de DIO ya que parece que el selenio se suministra en prioridad para la adecuada síntesis de las DIO a expensas de otras selenoenzimas (8).
La ingesta insuficiente de selenio puede afectar negativamente la actividad de varias enzimas sensibles al selenio, incluyendo las glutatión peroxidasas (GPx1 y GPx3), iodotironina deiodinasas, selenoproteínas W, y metionina-R-sulfóxido reductasa B1 (MsrB1). Incluso cuando es severa, la deficiencia aislada de selenio usualmente no deriva en una enfermedad clínica evidente. Sin embargo, a comparación de los sujetos con un estatus adecuado de selenio, los individuos con deficiencia de selenio podrían ser más susceptibles a estrés fisiológico adicional (41). La deficiencia prolongada de selenio puede contribuir a las enfermedades de Keshan y Kashin-Beck (véase abajo).
Se ha reportado deficiencia de selenio en pacientes crónicamente enfermos que recibieron nutrición parenteral total (NPT) sin selenio añadido por periodos de tiempo prolongados. Debilidad muscular, pérdida muscular, y cardiomiopatía (inflamación y daño al músculo cardíaco) han sido observados en estos pacientes. Hoy en día, las soluciones de NPT son rutinariamente suplementadas con selenio. El riesgo de una deficiencia de selenio puede incrementarse después de una cirugía bariátrica o en condiciones gastrointestinales severas, como la enfermedad de Crohn. Algunas dietas medicas especializadas como aquellas usadas para tratar ciertos desordenes metabólicos, incluyendo la fenilcetonuria, homocistinuria, y la enfermedad de la orina con olor a jarabe de arce, necesitan ser suplementadas con selenio para asegurar un estatus óptimo de selenio en los pacientes (42).
La enfermedad de Keshan es una forma fatal de cardiomiopatía dilatada que fue descrita primero en niños y mujeres jóvenes de una región en China con deficiencia de selenio. La forma aguda de esta enfermedad está caracterizada por la aparición repentina de insuficiencia cardíaca, mientras que la forma crónica resulta en un agrandamiento cardíaco de moderado a severo con grados variables de deficiencia cardíaca (43). La incidencia de la enfermedad de Keshan se asocia estrechamente con ingestas dietarías muy bajas de selenio y con un pobre estado nutricional del selenio. Se encontró que la suplementación con selenio (en la forma de selenito de sodio; Na2SeO3) protegió a las personas de desarrollar la enfermedad de Keshan pero no pudo revertir el daño al musculo cardiaco una vez que había ocurrido (43). Un reciente estudio de caso y control reporto que la actividad de la glutatión peroxidasa 1 sensible al selenio (GPx1) era significativamente menor en pacientes con Keshan en comparación a individuos saludables. Curiosamente, un polimorfismo de GPX1 específico que resulta en una transición de prolina a leucina en la posición 198 (Pro198Leu) está asociado con una reducción en la actividad de la GPx1 y fue encontrado más prevalente en los pacientes con Keshan. Este polimorfismo del GPX1 podría conferir una susceptibilidad mayor a la enfermedad de Keshan en portadores con un estatus nutricional de selenio bajo (44).
Mientras que la deficiencia de selenio es un factor etiológico fundamental de la enfermedad de Keshan, las variaciones estacionales y anuales en su ocurrencia sugieren que hay otros factores, especialmente un agente infeccioso podría estar involucrado además de la deficiencia de selenio (45). El Coxsackievirus B3 es un tipo de virus que ha sido aislado de pacientes con Keshan, y los estudios en animales han mostrado que este virus es capaz de causar una inflamación del corazón (miocarditis) en ratones deficientes de selenio. Los estudios en ratones también indicaron que el estrés oxidativo inducido por la deficiencia de selenio pudo resultar en cambios en el genoma viral; estos siendo capaces de convertir a una cepa relativamente inocua de coxsackievirus B3 en una cepa causante de miocarditis (43). Aunque aún sin ser probado en la enfermedad de Keshan, es posible que la deficiencia de selenio pueda incrementar la virulencia de los virus con el potencial de invadir y dañar el musculo cardíaco (46).
La enfermedad de Kashin-Beck (KBD) es otra condición endémica que afecta un estimado de 2.5 millones de personas en el Tíbet, norte y centro de China, Corea del Norte, y Sureste de Siberia (47). La KBD se caracteriza por una degeneración del cartílago articular entre las articulaciones (osteoartritis) que puede resultar en deformidades de las articulaciones y enanismo en las formas más severas de la enfermedad. La enfermedad afecta niños tan jóvenes como de dos años de edad. Al igual que con la enfermedad de Keshan, la KBD es prevalente en provincias deficientes de selenio y así generalmente afecta personas con ingestas muy bajas de selenio (47). Estudios recientes han sugerido que la susceptibilidad incrementada de KBD es las poblaciones deficientes de selenio podría resultar de una protección antioxidante reducida asociada con polimorfismos en los genes GPX (48, 49). A pesar de todo, la etiología parece ser multifactorial, ya que un cierto número de otros factores causantes han sido sugeridos para la KBD, incluyendo toxinas fúngicas en granos, deficiencia de yodo, y agua potable contaminada (43).
Meta-análisis recientes de unos pocos ensayos de menor magnitud y de estudios de cohorte prospectivos han indicado que el mejoramiento del estatus nutricional del selenio en niños que viven en áreas endémicas puede ayudar a reducir la incidencia de KBD (50). También, existe evidencia limitada que sugiere que la suplementación con selenio podría ser útil en el tratamiento de pacientes con KBD. Un meta-análisis de 10 ensayos controlados aleatorios reportó un incremento significante en la tasa de reparación de lesiones oseas en niños con KBD suplementados con selenito de sodio por al menos un año (51). Ensayos más amplios de mejor calidad son necesarios para evaluar si la suplementación con selenio podría resultar en remisión de la enfermedad.
La ingesta diaria recomendada (IDR) para el selenio fueron revisadas por última vez en el 2000 por la Junta de Nutrición y Alimentos (JNA) del Instituto de Medicina de los EE.UU. La IDR más reciente está basada en el requerimiento estimado promedio (REP) necesario para maximizar la actividad de la enzima antioxidante glutatión peroxidasa (GPx) en el plasma (52).
| Etapa de la Vida | Edad | Machos (μg/día) | Hembras (μg/día) |
|---|---|---|---|
| Infantes | 0-6 meses | 15 (IA) | 15 (IA) |
| Infantes | 7-12 meses | 20 (IA) | 20 (IA) |
| Niños | 1-3 años | 20 | 20 |
| Niños | 4-8 años | 30 | 30 |
| Niños | 9-13 años | 40 | 40 |
| Adolescentes | 14-18 años | 55 | 55 |
| Adultos | 19 años y más | 55 | 55 |
| Embarazo | Todas las edades | - | 60 |
| Período de lactancia | Todas las edades | - | 70 |
Debe notarse que la tercera Encuesta Nacional de Salud y Examinación de Nutrición (NHANES III) reportó que más del 99% de los participantes estadounidenses tuvieron concentraciones de selenio en el suero consistentes con los requerimientos de selenio requeridos (52), sugiriendo que la suplementación con selenio no es necesaria para los americanos.
Ha habido investigación considerable sobre el efecto de la suplementación con selenio en la incidencia de cáncer en animales. Mas de dos tercios de los más de 100 estudios publicados en 20 modelos diferentes de animales de canceres espontáneos, virales e inducidos químicamente encontró que la suplementación con selenio (a por lo menos niveles de ingesta adecuados) reduce significativamente la incidencia de tumores, especialmente en comparación a dietas deficientes de selenio (53). Evidencia de los efectos inhibidores de cáncer del selenio ha provisto de una sólida justificación para la investigación de asociaciones potenciales entre la ingesta de selenio y el riesgo de cáncer en humanos.
La mayoría de la evidencia epidemiológica temprana proveniente de estudios caso-control anidados y de caso y control sugirieron tanto asociaciones nulas como inversas entre la exposición al selenio y el riesgo de canceres de sitio especifico (54). Marcadores de la exposición al selenio incluyen el contenido de selenio sanguíneo y de las uñas del pie, como también la actividad de glutatión peroxidasa (GPx) en el plasma. Sin embargo, no está claro si reflejan adecuadamente la exposición al selenio proveniente de fuentes dietarías y suplementarias (véase Fuentes) o la distribución de selenio en los tejidos y órganos que podrían ser afectados por el cáncer. En el estudio prospectivo Danish Prospective Diet, Cancer, and Health, que dio seguimiento a más de 57,000 hombres y mujeres por 14 años, se encontró que el riesgo de cáncer rectal era 42% mayor en fumadores actuales en comparación a los no fumadores. Ninguna diferencia entre fumadores y no fumadores con respecto a las ingestas suplementarias y dietarías de micronutrientes antioxidantes, incluyendo selenio, que contribuyera a la asociación entre el fumar y el cáncer rectal fue encontrada (55). Sin embargo, debido a que estudios consistentemente han reportado bajas concentraciones de selenio en la sangre y bajas actividades de la GPx en los fumadores en comparación a los no fumadores (revisado en 56), el estimado de las ingestas de selenio podría no ser un marcador fidedigno de la exposición al selenio en esta población. También, las formas químicas del selenio encontradas en los alimentos son variadas (véase Fuentes) y pueden tener efectos biológicos y toxicológicos muy diferentes (57, 58).
Una revisión Cochrane reciente incluyó 55 estudios basados en la observación completados — mayormente con un diseño de caso-control anidado — publicados durante tres décadas (54). Un meta-análisis de 16 de estos estudios basados en la observación, incluyendo más de 144,000 individuos, reportó que el estatus de selenio más alto frente al más bajo estaba asociado con un riesgo 31% más bajo de cáncer en cualquier sitio y un riesgo 40% más bajo de mortalidad relacionada al cáncer. Un riesgo significantemente menor fue reportado para el cáncer de vejiga (5 estudios) y el cáncer de próstata (17 estudios); sin embargo, el estatus alto de selenio no estuvo inversamente relacionado con los riesgos de cáncer de seno (8 estudios), cáncer pulmonar (12 estudios), cáncer colorrectal (5 estudios), y cáncer gástrico (5 estudios) (54). Otro meta-análisis de 16 estudios basados en la observación reportó una relación inversa entre el cáncer de seno y las concentraciones de selenio en el suero (59). Las diferencias en género en la susceptibilidad al cáncer han sido reportadas en algunos estudios, aunque evidencia consistente de efectos diferentes en hombres y mujeres parece ser carente.
Variaciones de un solo nucleótido (polimorfismos) en la secuencia de los genes pueden modificar el nivel de expresión de genes y la estabilidad y actividad de las proteínas sintetizadas. Por ejemplo, una transición de prolina a leucina causada por un polimorfismo especifico en el gen GPX1 (rs1050450 C>T) está asociada con la actividad enzimática reducida de la GPx1. La descripción de varios polimorfismos en los genes que codifican para las selenoproteínas ha llevado a la evolución de posibles asociaciones con el estatus de selenio y la incidencia de cáncer. Notablemente, ciertos polimorfismos en los genes que codifican para las selenoproteínas han sido asociados con riesgos incrementados de canceres colorrectales y gástricos (revisado en 60). Adicionalmente, un cierto número de estudios han investigado el efecto de los polimorfismos de la selenoproteína en la relación entre el estatus del selenio y el riesgo de cáncer de próstata. Un estudio caso-control anidado dentro de la cohorte EPIC-Heidelberg ha combinado la genotipificación para varias variantes de la selenoproteína con marcadores del estatus del selenio (61). Brevemente, el estudio mostró que un polimorfismo del gen GPX1 (rs1050450 C>T) afectó la asociación entre las concentraciones de selenio y el riesgo de cáncer de próstata. Específicamente, se encontró que las concentraciones de selenio estaban inversamente asociadas con el riesgo de cáncer de próstata solo entre portadores del alelo GPX1. Variantes adicionales en los genes de la selenoproteína podrían mitigar el efecto del estatus del selenio en el riesgo de cáncer de próstata (62, 63). En otro estudio caso-control anidado dentro del Physicians’ Health Study (PHS), se encontró que los individuos en el cuartil más alto de las concentraciones de selenio frente aquellos en el cuartil más bajo tenían un riesgo reducido de mortalidad relacionada al cáncer de próstata a menos que portaran una variante especifica en el gen de la selenoproteína de 15kDa (SEP15 rs561104 G>A) (64). Mas investigación es necesaria para desentrañar aún más los mecanismos que subyacen la influencia de las interacciones genes-dieta en el riesgo de desarrollar cáncer.
Estudios basados en la comunidad: Un ensayo de intervención muy temprano de la suplementación con selenio se llevó a cabo en China entre una población general de 130,471 individuos viviendo en una zona con alto riesgo de infección por hepatitis B viral y cáncer de hígado. El ensayo proveyó sal de mesa enriquecida con selenito de sodio a la población de uno de los municipios (20,847 personas) usando a otros cuatro municipios como controles. Durante un periodo de seguimiento de ocho años, la incidencia promedio de cáncer de hígado se redujo en un 35% en la población enriquecida con selenio, mientras que no se encontró ninguna reducción en las poblaciones de control. En un ensayo clínico en la misma región, 226 individuos con evidencia de infección por hepatitis B crónica fueron suplementados diariamente con 200 μg de selenio en la forma de tabletas de levadura enriquecidas con selenio o con un placebo de tabletas de levadura. Durante el periodo de seguimiento de 4 años, 7 de 113 de los individuos que tomaban el placebo desarrollaron cáncer hepático primario, mientras que ninguno de los 113 sujetos suplementados con selenio desarrolló cáncer de hígado (65).
Ensayos controlados aleatorios: El estudio doble ciego, controlado con placebo para la Prevención Nutricional del Cáncer (NPC) en 1,312 adultos mayores con historial de cáncer de piel del tipo no melanoma, encontró que la suplementación con 200 μg/día de levadura enriquecida con selenio (levadura selenizada) por un promedio de 7.4 años resulto en una disminución del 52% en la incidencia de cáncer de próstata en hombres (revisado en 66). El efecto protector de la suplementación con selenio fue mayor en los hombres con niveles basales del plasma más bajos de selenio y del antígeno prostático específico (APE). Una incidencia reducida de cáncer total, pulmonar, y colorrectal fue asociada con la suplementación de 200 μg/día (67) pero no con 400 μg/día de levadura enriquecida con selenio (68). Además, la suplementación con selenio incremento el riesgo de un tipo de cáncer de piel (carcinoma celular escamoso) en un 25%. Un ensayo aleatorio de intervención, controlado con placebo de mayor escala (estudio SELECT) en más de 35,000 hombres de mediana edad y repletos de selenio, a los cuales se les asignó aleatoriamente una suplementación con selenio (en la forma de selenometionina, 200 μg/día) y/o vitamina E, fue detenido debido a preocupaciones con respecto a un riesgo incrementado de diabetes tipo 2 con el selenio y un riesgo incrementado de cáncer de próstata con la vitamina E (69, 70). Además, la suplementación con selenio, únicamente o junto con la vitamina E, no mostro ningún beneficio con respecto al riesgo de canceres de próstata, pulmón, o colorrectal después de un seguimiento de 5.5 años (71, 72). La carencia de un efecto beneficial de la suplementación con selenio fue apoyada en un reciente meta-análisis de ensayos controlados aleatorios (54).
Los niveles de actividad bajos de las selenoenzimas, glutatión peroxidasas (GPx), han sido reportados en enfermedades relacionadas al estrés oxidativo, incluyendo las enfermedades cardiovasculares (ECV) (73). En teoría, el mantenimiento de un estatus óptimo de selenio tiene el potencial de proteger contra el estrés oxidativo (incluyendo la peroxidación lipídica) y podría eventualmente prevenir la inflamación crónica y los trastornos cardiovasculares. Sin embargo, análisis de datos de corte transversal provenientes de 13,887 adultos estadounidenses incluidos en la Tercera Encuesta Nacional de Salud y Examinación de Nutrición (NHANES III, 1988-1994) fallaron en mostrar alguna asociación significante entre las concentraciones de selenio y la mortalidad por ECV, enfermedad coronaria cardiaca (ECC), o por accidente cerebrovascular (74). Además, mientras que individuos con insuficiencia renal están en un riesgo más alto de desarrollar ECC en comparación con aquellos con una función renal normal, tal riesgo no se encontró que fuese mayor con bajas en lugar de concentraciones normales de selenio en el suero (≤98 ng/mL vs. >98 ng/mL) (75).
Un estudio de corte transversal basado en los datos de NHANES 2003-2004 proveniente de 2,638 participantes de 40 años y más encontró que el riesgo de presión sanguínea alta (hipertensión), un factor contribuyente principal para las ECV era 73% más alto en individuos en el quintil más alto versus aquellos en el quintil más bajo de las concentraciones de selenio del suero (≥150 ng/mL vs. <122 ng/mL) (76). Sin embargo, una reciente revisión sistemática de la literatura falló en encontrar evidencia suficiente para apoyar alguna relación entre las concentraciones de selenio en el suero y la hipertensión (77). Además, unos pocos estudios basados en la observación han también reportado asociaciones entre el estatus normal a alto del selenio y los niveles de lípidos elevados en el suero en las poblaciones repletas de selenio, especulando que el selenio podría interferir con el metabolismo de las grasas y adversamente afectar la salud cardiovascular (78, 79). Hoy en día, ensayos controlados aleatorios no han provisto de resultados consistentes con respecto al efecto de la suplementación con selenio en los niveles lipídicos ni tampoco han demostrado algún beneficio cardiovascular del selenio en los individuos con ingestas óptimas o subóptimas de selenio (80).
La deficiencia de selenio ha sido asociada con una inmunidad deteriorada e inflamación crónica (81). Una cantidad considerable de investigación conducida en cultivo de células y modelos animales indica que el selenio juega papeles esenciales en la regulación de la migración, proliferación, diferenciación, activación y función optima de las células inmunes, influenciando así la inmunidad innata, la producción de anticuerpos dependientes de células B y la inmunidad mediada por células T (revisado en 82). Evidencia reciente en el papel del selenio y selenoproteínas en la producción de mediadores lipídicos (llamados eicosanoides) involucrados en las respuestas inflamatorias sugiere que la suplementación con selenio podría mitigar las respuestas inflamatorias disfuncionales que contribuyen a la patogénesis de muchas condiciones de salud crónicas (83). Hoy en día, ensayos controlados aleatorios son necesarios para evaluar los beneficios potenciales de la suplementación con selenio en los desórdenes inflamatorios, tales como el asma (84) y la enfermedad intestinal inflamatoria (85).
En áreas de malnutrición generalizada, las deficiencias de micronutrientes (incluyendo el selenio) son comunes en los individuos infectados con el virus de inmunodeficiencia humana (VIH) que causa el síndrome de inmunodeficiencia adquirida (SIDA). Antes de que la terapia antirretroviral (TAR) fuese estándar para el tratamiento del VIH, estudios basados en la observación habían consistentemente reportado asociaciones entre las concentraciones bajas de selenio en el suero y la infección de VIH en sujetos bien nutridos (86). El estatus pobre de selenio ha sido ligado a riesgos incrementados de cardiomiopatía dilatada y mortalidad en niños y adultos infectados con VIH, como también transmisión de VIH de madre a hijo y mortalidad perinatal (revisado en 87). Estudios de laboratorio tempranos han sugerido que el VIH podría interrumpir las defensas antioxidantes normales en las células T infectadas al reducir los niveles de selenoproteínas, es decir, tiorredoxina reductasas y glutatión peroxidasas (87). Curiosamente, un estudio de corte transversal encontró que los individuos VIH-seropositivos que recibían TAR por más de dos años tenían cargas virales indetectables en el plasma, conteos de linfocitos T CD4 más altos, y concentraciones de selenio adecuadas en el suero en comparación a sujetos no tratados previamente con TAR (88). Debido a que la actividad antioxidante de las selenoproteínas puede interferir con la replicación viral de las células inmunes infectadas con VIH (89, 90), se ha sugerido que la suplementación con selenio podría servir como un potencial adyuvante de la TAR para los pacientes con VIH.
Unos pocos ensayos de la suplementación con selenio en individuos infectados con VIH han sido conducidos. Un ensayo aleatorio, doble ciego, controlado con placebo en 186 adultos VIH positivos inicialmente encontró que la suplementación con 200 μg/día de selenio por dos años significativamente disminuyo la tasa de admisiones al hospital (91). Otro ensayo aleatorio, doble ciego, controlado con placebo en 174 individuos VIH positivos reportó que la suplementación con 200 μg/día de selenio (en la forma de levadura enriquecida con selenio) por nueve meses incremento las concentraciones de selenio del suero, mejoró el conteo de linfocitos T CD4, y previno cualquier progresión de la carga viral del VIH (92). En un tercer ensayo doble ciego en Tanzania, 913 mujeres embarazadas de entre 12 y 27 semanas de gestación fueron aleatoriamente asignadas a recibir 200 μg/día de selenio (como selenometionina) o un placebo hasta los seis meses después del nacimiento. La suplementación con selenio no tuvo efecto en el conteo de células T CD4, CD8, y CD3 y en la carga viral del VIH, pero significantemente disminuyó el riesgo de diarrea aguda o persistente (93, 94). Además, el riesgo de muerte entre las seis semanas y seis meses después del parto fue significantemente reducido en infantes y madres suplementados con selenio en comparación al placebo (94).
Un reciente ensayo de cuatro brazos en Botswana aleatoriamente asigno 878 adultos VIH positivos en etapa temprana de la infección a recibir tanto un tratamiento con placebo, multivitaminas (vitamina B, C, y E), 200 μg/día de selenio, o ambos multivitaminas y selenio por 24 meses (95). A diferencia del selenio solo, la suplementación con multivitaminas (con o sin selenio) disminuyó el riesgo de declive inmunológico al significantemente incrementar el tiempo antes de que la iniciación de TAR fuese necesaria (es decir, cuando el conteo de células T CD4 cayera por debajo de las 251 células/mcL) en comparación al placebo. En el estudio, un resultado combinado de (1) conteo de células T CD4 cayendo por debajo de las 251 células/mcL; (2) la ocurrencia de condiciones que definen el SIDA; y (3) muerte relacionada al SIDA — cualquiera que ocurriese primero — fue usado para evaluar la progresión de la enfermedad en los diferentes brazos del tratamiento. En comparación al placebo, hubo un periodo de tiempo más largo a partir de la aleatorización hasta la fecha del resultado compuesto en individuos suplementados con multivitaminas más selenio, pero no en aquellos que recibieron multivitaminas o selenio únicamente (95). Más investigación es necesaria para replicar estos resultados preliminares, especialmente en condiciones y comunidades donde la malnutrición acelera la progresión de la infección del VIH y el acceso a la terapia retroviral pueda ser limitada.
El síndrome de respuesta inflamatoria sistémica (SIRS) resulta de una respuesta inflamatoria sistémica que puede ocurrir debido a una infección (sepsis) (96). La sepsis severa y el choque séptico — definido como una baja presión sanguínea inducida por la sepsis persistente — están asociados con tasas elevadas de mortalidad en pacientes en estado crítico (96, 97). Debido a que las respuestas inflamatorias sistémicas involucran estrés oxidativo excesivo, se ha sugerido que el proporcionar nutrientes antioxidantes como el selenio puede mejorar el resultado de pacientes en estado crítico en unidades de cuidado intensivo. Dos meta-análisis recientes de ensayos controlados aleatorios encontró que la suplementación intravenosa con selenio (como selenito de sodio) en pacientes en estado crítico padeciendo de SIRS, sepsis, o choque séptico resulto en un riesgo significantemente reducido de mortalidad en un 17% a 27% (98, 99). Más ensayos son necesarios para identificar el programa apropiado de la administración de selenio (en términos de dosis, ruta, y duración del tratamiento) y para evaluar los resultados adicionales (p. ej., complicaciones infecciosas y duración de la estadía en el hospital).
La tiroiditis de Hashimoto (TH; tiroiditis autoinmune crónica) es una enfermedad autoinmune caracterizada por la infiltración de células T en la glándula tiroides y autoanticuerpos circulantes (predominantemente contra la peroxidasa tiroidea), causando inflamación prolongada, daño a los tejidos, e hipotiroidismo (8). Mientras que la función de la glándula tiroides de los individuos saludables esta usualmente protegida de variaciones en el suministro del selenio, la deficiencia de selenio y los polimorfismos genéticos que afectan la actividad de las selenoproteínas podrían ser factores contribuyentes potenciales para las enfermedades autoinmunes de la tiroides. Una reciente revisión sistemática (100) identificó cuatro ensayos controlados aleatorios que evaluaron el efecto de la suplementación con selenio como un tratamiento adyuvante para la terapia de remplazo de T4 (levotiroxina) en pacientes con TH (101-104). Mientras que tres de cuatro estudios sugirieron una reducción en los niveles de autoanticuerpos circulantes, ninguno de ellos proporciono información sobre si el selenio puede mejorar los síntomas relacionados a la salud y el estado de ánimo para permitir una dosis disminuida de levotiroxina. Otro ensayo controlado aleatorio encontró que la suplementación con selenio mejoró el bienestar de los pacientes afectados por otra enfermedad autoinmune de la tiroides que conduce al hipertiroidismo (enfermedad de Graves) (105). Los resultados de dos ensayos aleatorios, controlados con placebo en proceso — el ensayo CATALYST en pacientes con TH y el ensayo GRASS en pacientes con enfermedad de Graves — podrían proporcionar una visión en un efecto del selenio en el criterio de la calidad de vida específica a la tiroides e informar los criterios de toma de decisiones clínicas (106, 107).
Las fuentes alimenticias más ricas de selenio son las vísceras y los mariscos, seguidos de las carnes con músculo. El agua potable no se considera como una fuente significante de selenio en Norteamérica. Sin embargo, en áreas, donde los niveles altos de selenio en el suelo contribuyen al contenido de selenio del agua, niveles más altos de selenio pueden ser encontrados en pozos usados para el agua potable (108). En general, existe una amplia variación en el contenido de selenio de plantas y granos, especialmente debido a que algunas plantas incluyendo el ajo, las nueces de Brasil, y múltiples Brassica especies, tienden a acumular selenio ("acumuladores de selenio"), mientras otras asimilan selenio en menor grado ("no-acumuladores"). La asimilación de selenio por las plantas también depende del contenido de selenio del suelo. Las nueces de Brasil que crecen en áreas de Brasil con suelos ricos en selenio pueden proporcionar más de 100 μg de selenio en una nuez, mientras que aquellas que crecen en suelos carentes de selenio proporcionan 10 veces menos (109). En los EE.UU., los granos son una buena fuente de selenio, pero las frutas y verduras tienden a ser fuentes de selenio relativamente pobres.
Varias formas químicas (especies) de selenio se encuentran en acumuladores de selenio, incluyendo el selenato (selenio inorgánico), la selenometionina, la selenocisteína, la selenio-metil-selenocisteína, y la γ-glutamil-selenio-metil-selenocisteína. Aunque los últimos dos compuestos son predominantes en plantas de las familias Allium y Brassicaceae (las cuales incluyen a los ajos, cebollas, y brócoli), el trigo, otros granos (incluyendo las nueces de Brasil), y la soya son ricos en selenometionina y contienen pequeñas cantidades de selenocisteína y selenato. Se conoce menos acerca de las especies de selenio y la distribución en fuentes dietarías de origen animal. La nutrición animal y las condiciones de crecimiento ciertamente contribuyen a las especies de selenio formadas y sus cantidades relativas, y se asume que la ruta metabólica del selenio dietario en animales es similar a la de los humanos. La selenocisteína es predominantemente formada en animales que se alimentan de selenio inorgánico, mientras que la selenometionina se deriva de las fuentes dietarías de origen vegetal (revisado en 110).
En los EE.UU., el cuestionario nacional NHANES III reportó ingestas dietarías promedio oscilando entre los 100.5 μg/día y 158.5 μg/día en adultos de entre 19-50 años de edad (52). La Tabla 2 lista algunas fuentes alimenticias ricas en selenio y su contenido de selenio en microgramos (μg). Para más información en el contenido de selenio de alimentos específicos, revise la base de datos de composición de los alimentos de la USDA.
| Alimento | Porción | Selenio (μg) |
|---|---|---|
| Nueces de Brasil (de suelo rico en selenio) | 1 onza (6 unidades) | 543.5* |
| Atún (aleta amarilla, cocido, seco al calor) | 3 onzas | 92.0 |
| Ostras (Pacifico, crudas) | 3 onzas | 65.4 |
| Almejas (mixtas, cocidas, al vapor) | 3 onzas | 54.4 |
| Hipogloso (Atlántico y Pacífico, cocido, seco al calor) | 3 onzas | 47.1 |
| Camarón (cocido, al vapor) | 3 onzas | 42.1 |
| Salmón (Chinook, cocido, seco al calor) | 3 onzas | 39.8 |
| Fideos (de huevo, cocido, enriquecidos) | 1 taza | 38.2 |
| Cangrejo (reina, cocido, al vapor) | 3 onzas | 37.7 |
| Puerco (carne magra, lomo, cocido, rostizado) | 3 onzas | 32.5 |
| Carne de res (magra, bistec, cocida, a la parrilla) | 3 onzas | 30.6 |
| Pollo (carne blanca, cocida, rostizada) | 3 onzas | 25.8 |
| Arroz (integral, grano grande, cocido) | 1 taza | 19.1 |
| Semillas de girasol (secas) | ¼ taza | 18.6 |
| Pan de trigo entero | 2 rebandas | 16.4 |
| Leche (sin grasa o descremada) | 8 onzas fluidas (1 taza) | 7.6 |
| *Por encima del nivel máximo de ingesta tolerable (NM) de 400 μg/día. | ||
Los suplementos de selenio están disponibles en varias formas. El selenito y el selenato de sodio son formas inorgánicas del selenio. El selenato de sodio es absorbido casi completamente, pero se excreta una cantidad significativa en la orina antes de que pueda ser incorporado a las proteínas. El selenito de sodio sólo se absorbe en un 50% pero es retenido de mejor manera que el selenato una vez que es absorbido. La selenometionina es absorbida en aproximadamente un 90% (52); sin embargo, solo alrededor de un 34% puede ser actualmente convertida en selenometionina libre (111). La selenometionina y la levadura enriquecida con selenio, los cuales aportan principalmente selenometionina, también se encuentran disponibles como suplementos. El consumidor debería estar al tanto de que algunas formas de levaduras enriquecidas con selenio disponibles contienen levadura más formas de selenio principalmente inorgánicas.
Aunque tanto las formas orgánicas como las inorgánicas del selenio pueden ser metabolizadas a selenocisteína por el cuerpo y ser incorporadas en las selenoenzimas, podrían no contribuir igualmente al mantenimiento de un estatus de selenio adecuado. En ensayos de intervención, la suplementación con una forma orgánica de selenio (selenometionina) incremento más efectivamente las concentraciones de selenio en la sangre en comparación a la suplementación con formas inorgánicas (es decir, el selenito de sodio y selenato de sodio) (110). Sin embargo, las formas inorgánicas pueden incrementar la actividad de la glutatión peroxidasa (GPx) en el plasma más efectivamente que las formas orgánicas (revisado en 112). También se ha sugerido que la incorporación de selenometionina en lugar de metionina en las proteínas de los tejidos puede asegurar que el selenio esté disponible tras el recambio proteico (110).
Los alimentos enriquecidos con selenio han sido de interés para los científicos, especialmente debido a la insinuación de que algunas de las formas químicas del selenio producidas por las plantas podrían ser modificadores del riesgo de cáncer más potentes que aquellas disponibles actualmente en suplementos. Aunque no existe evidencia actual de los beneficios a largo plazo de la salud asociados con el consumo de alimentos enriquecidos con selenio, resultados de estudios en animales y ensayos de intervención sugieren que las fuentes basadas en proteína de selenio son más efectivas en incrementar la actividad de GPx que la levadura enriquecida con selenio y selenometionina (112). La fortificación de alimentos puede también representar una estrategia rentable para mejorar el estatus nutricional de selenio en poblaciones en riesgo de insuficiencia (113).
Aunque el selenio es requerido para la salud, altas dosis de selenio pueden ser toxicas. La toxicidad aguda y fatal ha ocurrido por la ingesta accidental o suicida de cantidades en gramos de selenio. Se reportó toxicidad por selenio clínicamente significativa en 13 individuos luego de que tomaran suplementos que contenían 27.3 mg (27,300 μg) por tableta debido a una falla de fabricación. La toxicidad crónica por selenio (selenosis) puede ocurrir con dosis de selenio más pequeñas por largos periodos de tiempo. Los síntomas de selenosis reportados con mayor frecuencia son la pérdida y fragilidad de uñas y cabello. Otros síntomas podrían incluir molestias gastrointestinales, erupciones cutáneas, aliento con olor a ajo, fatiga, irritabilidad, y trastornos neurológicos. En un área de China con una alta prevalencia de selenosis, los efectos tóxicos ocurrieron con una mayor frecuencia cuando las concentraciones de selenio sanguíneo alcanzaban un nivel correspondiente a una ingesta de 850 μg/día.
La Junta de Nutrición y Alimentos (JNA) del Instituto de Medicina de los EE.UU. estableció el nivel máximo de ingesta tolerable (NM) para el selenio en 400 μg/día para adultos basado en la prevención de la pérdida y fragilidad de uñas y cabello y de signos iniciales de la toxicidad crónica por selenio (52). El NM de 400 μg/día para los adultos incluyen tanto el selenio obtenido de alimentos como el selenio de suplementos (Tabla 3).
Unos pocos estudios han examinado la relación entre el estatus de selenio y la diabetes mellitus tipo 2. En el análisis de corte transversal de datos del NHANES III (1988-1994) provenientes de 8,876 participantes adultos, el quintil más alto frente al más bajo de las concentraciones de selenio en el suero (≥137 ng/mL vs. <111 ng/mL) se asoció con un riesgo incrementado de diabetes tipo 2 (114). Análisis de datos de 917 participantes (≥40 años de edad) del NHANES 2003-2004 también indicaron un incremento en la prevalencia de diabetes tipo 2 en el cuartil más alto de las concentraciones de selenio en el suero frente al cuartil más bajo (≥147 ngm/L vs. <124 ng/mL). Los individuos en el cuartil más alto frente aquellos en el más bajo de las concentraciones de selenio del suero tuvieron también niveles más altos de glucosa en el plasma y hemoglobina glicosilada, sugiriendo un control glicémico pobre (115). El estudio aleatorio, doble ciego, controlado con placebo en 1,312 participantes en el ensayo de Prevención Nutricional del Cáncer (NPC) encontró que la suplementación con selenio (200 μg/día; seguimiento promedio de 7.7 años) significantemente incremento el riesgo de diabetes tipo 2 en participantes en el tercil más alto de las concentraciones de selenio basales del plasma (116). Además, en el ensayo SELECT (Selenium and Vitamin E Cancer Prevention Trial), más casos de diabetes tipo 2 fueron encontrados en el grupo de selenio (200 μg/día; seguimiento medio de 5.5 años) en comparación al grupo de placebo, pero esto solo fue una tendencia y no fue estadísticamente significante (72).
En la actualidad, los mecanismos detrás de estas observaciones no son bien entendidos. Un incremento en la sensibilidad a la insulina ha sido reportado en individuos con deficiencia congénita (de nacimiento) de la mayoría de selenoproteínas (117). Los resultados de varios estudios en animales también indicaron que la suplementación con selenio y las selenoproteínas pueden interferir con la acción de la insulina y con la homeostasis de la glucosa (revisado en 118). Por otra parte, estudios recientes han encontrado que el metabolismo deteriorado de la glucosa en pacientes con diabetes tipo 2 puede afectar la expresión de SEPP1 y la homeostasis del selenio (11, 119, 120). Mientras que más investigación es necesaria para completamente entender la interacción entre el metabolismo de los carbohidratos y la homeostasis del selenio, el uso de suplementos de selenio es considerado menos propenso a incrementar el riesgo de diabetes tipo 2 en individuos saludables, pero debe ser desalentado en aquellos con un estatus alto de selenio y/o en un riesgo incrementado de desarrollar diabetes tipo 2 (118).
En la actualidad, pocas interacciones entre el selenio y medicamentos han sido reportadas (121). Por ejemplo, el medicamento anticonvulsivo ácido valpróico y el agente quimioterapéutico cisplatino pueden disminuir las concentraciones de selenio circulante en sujetos tratados (122, 123). También se ha encontrado en estudios en animales que el selenito de sodio reduce la toxicidad del antibiótico nitrofuratoina y del herbicida paraquat (124).
Un ensayo aleatorizado controlado de tres años en 160 pacientes con enfermedad coronaria cardíaca (ECC) documentada y bajos niveles de lipoproteínas de alta densidad (HDL) encontró que una combinación de simvastatina (Zocor) y niacina incrementó los niveles subfracción de HDL2, inhibió la progresión de estenosis (estrechamiento) arterial coronaria, y disminuyó la frecuencia de eventos cardiovasculares (125). Sorprendentemente, cuando se tomó una combinación antioxidante (1,000 mg de vitamina C, 800 UI de vitamina E (d-α-tocoferol), 100 μg de selenio, y 25 mg de β-caroteno diariamente) junto a la combinación de simvastatina-niacina, los efectos protectores fueron disminuidos. Sin embargo, la contribución individual del selenio no se puede determinar, y otros estudios han reportado que las vitaminas antioxidantes solas pudieron interferir con la acción de los medicamentos que elevan los HDL incluyendo las estatinas (126).
Se estima que la dieta Americana promedio aporta cerca de 100 μg/día de selenio, una cantidad bastante superior a la actual IDR (55 μg/día) y que parece ser suficiente para optimizar la actividad de la glutatión peroxidasa (GPx) celular y del plasma. Un reciente estudio controlado aleatorio de 10 semanas en adultos británicos saludables (edades, 50-64 años) estimó que una ingesta total de selenio de alrededor de 105 μg/día era requerida para maximizar las concentraciones de selenoproteínas P (SEPP1), otro biomarcador útil del estatus del selenio (127). Sin embargo, un ensayo similar en una cohorte americana con altas concentraciones basales de selenio en el plasma no encontró efecto alguno de la suplementación con selenio en las concentraciones de SEPP1 (128). Mientras que la cantidad de selenio varía considerablemente en los suplementos multivitamínicos/minerales (MVM), estos suplementos raramente aportan más que el Valor Diario (VD) de 70 μg. Consumir una dieta variada y tomar un suplemento MVM diariamente debiera aportar selenio suficiente a la mayoría de las personas en los EE.UU. y ayudaría a mejorar el estatus de selenio en las poblaciones con ingestas menores de selenio fuera de los EE.UU.
En la actualidad, el efecto de la suplementación con selenio en el riesgo de cáncer no es lo suficiente clara para apoyar una recomendación general para un suplemento extra de selenio, especialmente en hombres con concentraciones de suero consistentes con ingestas adecuadas de selenio. El ensayo SELECT encontró que 200 μg/día de selenio suplementario no disminuyó el riesgo de cáncer de próstata en hombres (72), refutando los resultados del ensayo NPC (véase Cáncer). Otro estudio multicéntrico, aleatorio, doble ciego, controlado con placebo (the Negative Biopsy Trial) en 699 hombres en alto riesgo de cáncer de próstata no encontró efecto alguno de 200 μg/día o 400 μg/día de selenio en el riesgo de cáncer de próstata durante un seguimiento promedio de 36 meses (129). Además, debido a que evidencia actual sugiere una relación en forma de U entre el estatus de selenio y el riesgo de cáncer de próstata (130), los hombres debiesen evitar tomar selenio suplementario que sobrepase 200 μg/día.
En la actualidad, no existe evidencia clínica que demuestre que la suplementación con selenio por arriba de los niveles recomendados disminuya el riesgo de cáncer de seno aunque algunos, pero no todos, estudios basados en la observación han encontrado una relación inversa entre el estatus del selenio y el cáncer de seno en mujeres (59).
El envejecimiento no se ha asociado con cambios significativos en el requerimiento de selenio. Un ensayo aleatorio, doble ciego, controlado con placebo de cinco años en personas danesas mayores saludables (edades de inclusión, 60-74 años) encontró que la suplementación con selenio (100-300 μg/día) tuvo poco a no impacto en los niveles de enzimas antioxidantes, incluyendo GPx (131).
Originalmente escrito en 2001 por:
Jane Higdon, Ph.D.
Instituto Linus Pauling
Universidad Estatal de Oregon
Actualizado en Octubre de 2003 por:
Jane Higdon, Ph.D.
Instituto Linus Pauling
Universidad Estatal de Oregon
Actualizado en Noviembre de 2007 por:
Victoria J. Drake, Ph.D.
Instituto Linus Pauling
Universidad Estatal de Oregon
Actualizado en Noviembre de 2014 por:
Barbara Delage, Ph.D.
Instituto Linus Pauling
Universidad Estatal de Oregon
Revisado en Junio de 2015 por:
Petra A. Tsuji, Ph.D., M.P.H.
Profesor Asistente
Departamento de Ciencias Biológicas
Universidad de Towson
Traducido al Español en 2017 por:
Silvia Vazquez Lima
Instituto Linus Pauling
Universidad Estatal de Oregon
Originalmente traducido al español en 2012 por Guillermo Sandoval y editado por Andrew Quest (Ph.D.) y Lisette Leyton (Ph.D.), todos provenientes de la Universidad de Chile. Estos esfuerzos fueron patrocinados por el projecto Anillo #ACT1111, CONICYT-Chile, programa PIA.
Derechos de autoría 2001-2024 Instituto Linus Pauling
1. Rayman MP. The importance of selenium to human health. Lancet. 2000;356(9225):233-241. (PubMed)
2. Mangiapane E, Pessione A, Pessione E. Selenium and selenoproteins: an overview on different biological systems. Curr Protein Pept Sci. 2014;15(6):598-607. (PubMed)
3. Mariotti M, Ridge PG, Zhang Y, et al. Composition and evolution of the vertebrate and mammalian selenoproteomes. PLoS One. 2012;7(3):e33066. (PubMed)
4. Terry EN, Diamond JA. Selenium. In: Erdman Jr J, Macdonald J, Zeisel S, eds. Present Knowledge in Nutrition. 10th ed. Ames: Wiley-Blackwell; 2012:568-585.
5. Boitani C, Puglisi R. Selenium, a key element in spermatogenesis and male fertility. Adv Exp Med Biol. 2008;636:65-73. (PubMed)
6. Arner ES. Focus on mammalian thioredoxin reductases--important selenoproteins with versatile functions. Biochim Biophys Acta. 2009;1790(6):495-526. (PubMed)
7. Lu J, Holmgren A. The thioredoxin antioxidant system. Free Radic Biol Med. 2014;66:75-87. (PubMed)
8. Schomburg L. Selenium, selenoproteins and the thyroid gland: interactions in health and disease. Nat Rev Endocrinol. 2012;8(3):160-171. (PubMed)
9. Hill KE, Wu S, Motley AK, et al. Production of selenoprotein P (Sepp1) by hepatocytes is central to selenium homeostasis. J Biol Chem. 2012;287(48):40414-40424. (PubMed)
10. Olson GE, Winfrey VP, Hill KE, Burk RF. Megalin mediates selenoprotein P uptake by kidney proximal tubule epithelial cells. J Biol Chem. 2008;283(11):6854-6860. (PubMed)
11. Misu H, Takamura T, Takayama H, et al. A liver-derived secretory protein, selenoprotein P, causes insulin resistance. Cell Metab. 2010;12(5):483-495. (PubMed)
12. Whanger PD. Selenoprotein expression and function-selenoprotein W. Biochim Biophys Acta. 2009;1790(11):1448-1452. (PubMed)
13. Jeong D, Kim TS, Chung YW, Lee BJ, Kim IY. Selenoprotein W is a glutathione-dependent antioxidant in vivo. FEBS Lett. 2002;517(1-3):225-228. (PubMed)
14. Reeves MA, Hoffmann PR. The human selenoproteome: recent insights into functions and regulation. Cell Mol Life Sci. 2009;66(15):2457-2478. (PubMed)
15. Reszka E, Jablonska E, Gromadzinska J, Wasowicz W. Relevance of selenoprotein transcripts for selenium status in humans. Genes Nutr. 2012;7(2):127-137. (PubMed)
16. Chung YW, Jeong D, Noh OJ, et al. Antioxidative role of selenoprotein W in oxidant-induced mouse embryonic neuronal cell death. Mol Cells. 2009;27(5):609-613. (PubMed)
17. Jeon YH, Park YH, Kwon JH, Lee JH, Kim IY. Inhibition of 14-3-3 binding to Rictor of mTORC2 for Akt phosphorylation at Ser473 is regulated by selenoprotein W. Biochim Biophys Acta. 2013;1833(10):2135-2142. (PubMed)
18. Jeon YH, Park YH, Lee JH, Hong JH, Kim IY. Selenoprotein W enhances skeletal muscle differentiation by inhibiting TAZ binding to 14-3-3 protein. Biochim Biophys Acta. 2014;1843(7):1356-1364. (PubMed)
19. Alkan Z, Duong FL, Hawkes WC. Selenoprotein W controls epidermal growth factor receptor surface expression, activation and degradation via receptor ubiquitination. Biochim Biophys Acta. 2015;1853(5):1087-1095. (PubMed)
20. Ganichkin OM, Xu XM, Carlson BA, et al. Structure and catalytic mechanism of eukaryotic selenocysteine synthase. J Biol Chem. 2008;283(9):5849-5865. (PubMed)
21. Lee BC, Peterfi Z, Hoffmann FW, et al. MsrB1 and MICALs regulate actin assembly and macrophage function via reversible stereoselective methionine oxidation. Mol Cell. 2013;51(3):397-404. (PubMed)
22. Kim HY. The methionine sulfoxide reduction system: selenium utilization and methionine sulfoxide reductase enzymes and their functions. Antioxid Redox Signal. 2013;19(9):958-969. (PubMed)
23. Kumaraswamy E, Malykh A, Korotkov KV, et al. Structure-expression relationships of the 15-kDa selenoprotein gene. Possible role of the protein in cancer etiology. J Biol Chem. 2000;275(45):35540-35547. (PubMed)
24. Korotkov KV, Kumaraswamy E, Zhou Y, Hatfield DL, Gladyshev VN. Association between the 15-kDa selenoprotein and UDP-glucose:glycoprotein glucosyltransferase in the endoplasmic reticulum of mammalian cells. J Biol Chem. 2001;276(18):15330-15336. (PubMed)
25. Labunskyy VM, Ferguson AD, Fomenko DE, Chelliah Y, Hatfield DL, Gladyshev VN. A novel cysteine-rich domain of Sep15 mediates the interaction with UDP-glucose:glycoprotein glucosyltransferase. J Biol Chem. 2005;280(45):37839-37845. (PubMed)
26. Labunskyy VM, Hatfield DL, Gladyshev VN. The Sep15 protein family: roles in disulfide bond formation and quality control in the endoplasmic reticulum. IUBMB Life. 2007;59(1):1-5. (PubMed)
27. Kasaikina MV, Fomenko DE, Labunskyy VM, et al. Roles of the 15-kDa selenoprotein (Sep15) in redox homeostasis and cataract development revealed by the analysis of Sep 15 knockout mice. J Biol Chem. 2011;286(38):33203-33212. (PubMed)
28. Hatfield DL, Tsuji PA, Carlson BA, Gladyshev VN. Selenium and selenocysteine: roles in cancer, health, and development. Trends Biochem Sci. 2014;39(3):112-120. (PubMed)
29. Curran JE, Jowett JB, Elliott KS, et al. Genetic variation in selenoprotein S influences inflammatory response. Nat Genet. 2005;37(11):1234-1241. (PubMed)
30. Santos LR, Duraes C, Mendes A, et al. A polymorphism in the promoter region of the selenoprotein S gene (SEPS1) contributes to Hashimoto's thyroiditis susceptibility. J Clin Endocrinol Metab. 2014;99(4):E719-723. (PubMed)
31. Alanne M, Kristiansson K, Auro K, et al. Variation in the selenoprotein S gene locus is associated with coronary heart disease and ischemic stroke in two independent Finnish cohorts. Hum Genet. 2007;122(3-4):355-365. (PubMed)
32. Cox AJ, Lehtinen AB, Xu J, et al. Polymorphisms in the Selenoprotein S gene and subclinical cardiovascular disease in the Diabetes Heart Study. Acta Diabetol. 2013;50(3):391-399. (PubMed)
33. Moses EK, Johnson MP, Tommerdal L, et al. Genetic association of preeclampsia to the inflammatory response gene SEPS1. Am J Obstet Gynecol. 2008;198(3):336 e331-335. (PubMed)
34. Shibata T, Arisawa T, Tahara T, et al. Selenoprotein S (SEPS1) gene -105G>A promoter polymorphism influences the susceptibility to gastric cancer in the Japanese population. BMC Gastroenterol. 2009;9:2. (PubMed)
35. Shchedrina VA, Zhang Y, Labunskyy VM, Hatfield DL, Gladyshev VN. Structure-function relations, physiological roles, and evolution of mammalian ER-resident selenoproteins. Antioxid Redox Signal. 2010;12(7):839-849. (PubMed)
36. Li X, Hill KE, Burk RF, May JM. Selenium spares ascorbate and alpha-tocopherol in cultured liver cell lines under oxidant stress. FEBS Lett. 2001;508(3):489-492. (PubMed)
37. May JM, Mendiratta S, Hill KE, Burk RF. Reduction of dehydroascorbate to ascorbate by the selenoenzyme thioredoxin reductase. J Biol Chem. 1997;272(36):22607-22610. (PubMed)
38. Murer SB, Aeberli I, Braegger CP, et al. Antioxidant supplements reduced oxidative stress and stabilized liver function tests but did not reduce inflammation in a randomized controlled trial in obese children and adolescents. J Nutr. 2014;144(2):193-201. (PubMed)
39. Schneider MJ, Fiering SN, Thai B, et al. Targeted disruption of the type 1 selenodeiodinase gene (Dio1) results in marked changes in thyroid hormone economy in mice. Endocrinology. 2006;147(1):580-589. (PubMed)
40. Hess SY. The impact of common micronutrient deficiencies on iodine and thyroid metabolism: the evidence from human studies. Best Pract Res Clin Endocrinol Metab. 2010;24(1):117-132. (PubMed)
41. Thomson CD. Assessment of requirements for selenium and adequacy of selenium status: a review. Eur J Clin Nutr. 2004;58(3):391-402. (PubMed)
42. Cooper A, Mones RL, Heird WC. Nutritional management of infants and children with specific diseases and other conditions. In: Ross AC, Caballero B, Cousins RJ, Tucker KL, Ziegler TR, eds. Modern Nutrition in Health and Disease. 11th ed. Baltimore: Lippincott Williams & Wilkins; 2012:988-1005.
43. Sunde RA. Selenium. In: Ross AC, Caballero B, Cousins RJ, Tucker KL, Ziegler TR, eds. Modern Nutrition in Health and Disease. 11th ed. Baltimore: Lippincott Williams & Wilkins; 2014:265-276.
44. Lei C, Niu X, Wei J, Zhu J, Zhu Y. Interaction of glutathione peroxidase-1 and selenium in endemic dilated cardiomyopathy. Clin Chim Acta. 2009;399(1-2):102-108. (PubMed)
45. Chen J. An original discovery: selenium deficiency and Keshan disease (an endemic heart disease). Asia Pac J Clin Nutr. 2012;21(3):320-326. (PubMed)
46. Harthill M. Review: micronutrient selenium deficiency influences evolution of some viral infectious diseases. Biol Trace Elem Res. 2011;143(3):1325-1336. (PubMed)
47. Stone R. Diseases. A medical mystery in middle China. Science. 2009;324(5933):1378-1381. (PubMed)
48. Du XH, Dai XX, Xia Song R, et al. SNP and mRNA expression for glutathione peroxidase 4 in Kashin-Beck disease. Br J Nutr. 2012;107(2):164-169. (PubMed)
49. Xiong YM, Mo XY, Zou XZ, et al. Association study between polymorphisms in selenoprotein genes and susceptibility to Kashin-Beck disease. Osteoarthritis Cartilage. 2010;18(6):817-824. (PubMed)
50. Zou K, Liu G, Wu T, Du L. Selenium for preventing Kashin-Beck osteoarthropathy in children: a meta-analysis. Osteoarthritis Cartilage. 2009;17(2):144-151. (PubMed)
51. Jirong Y, Huiyun P, Zhongzhe Y, et al. Sodium selenite for treatment of Kashin-Beck disease in children: a systematic review of randomised controlled trials. Osteoarthritis Cartilage. 2012;20(7):605-613. (PubMed)
52. Food and Nutrition Board, Institute of Medicine. Selenium. Dietary reference intakes for vitamin C, vitamin E, selenium, and carotenoids. Washington, D.C.: National Academy Press; 2000:284-324. (National Academy Press)
53. Combs GF, Jr., Gray WP. Chemopreventive agents: selenium. Pharmacol Ther. 1998;79(3):179-192. (PubMed)
54. Vinceti M, Dennert G, Crespi CM, et al. Selenium for preventing cancer. Cochrane Database Syst Rev. 2014;3:CD005195. (PubMed)
55. Hansen RD, Albieri V, Tjonneland A, Overvad K, Andersen KK, Raaschou-Nielsen O. Effects of smoking and antioxidant micronutrients on risk of colorectal cancer. Clin Gastroenterol Hepatol. 2013;11(4):406-415 e403. (PubMed)
56. Northrop-Clewes CA, Thurnham DI. Monitoring micronutrients in cigarette smokers. Clin Chim Acta. 2007;377(1-2):14-38. (PubMed)
57. Hazane-Puch F, Champelovier P, Arnaud J, et al. Long-term selenium supplementation in HaCaT cells: importance of chemical form for antagonist (protective versus toxic) activities. Biol Trace Elem Res. 2013;154(2):288-298. (PubMed)
58. Weekley CM, Harris HH. Which form is that? The importance of selenium speciation and metabolism in the prevention and treatment of disease. Chem Soc Rev. 2013;42(23):8870-8894. (PubMed)
59. Babaknejad N, Sayehmiri F, Sayehmiri K, et al. The relationship between selenium levels and breast cancer: a systematic review and meta-analysis. Biol Trace Elem Res. 2014;159(1-3):1-7. (PubMed)
60. Meplan C, Hesketh J. The influence of selenium and selenoprotein gene variants on colorectal cancer risk. Mutagenesis. 2012;27(2):177-186. (PubMed)
61. Steinbrecher A, Meplan C, Hesketh J, et al. Effects of selenium status and polymorphisms in selenoprotein genes on prostate cancer risk in a prospective study of European men. Cancer Epidemiol Biomarkers Prev. 2010;19(11):2958-2968. (PubMed)
62. Gerstenberger JP, Bauer SR, Van Blarigan EL, et al. Selenoprotein and antioxidant genes and the risk of high-grade prostate cancer and prostate cancer recurrence. Prostate. 2015;75(1):60-69. (PubMed)
63. Meplan C, Rohrmann S, Steinbrecher A, et al. Polymorphisms in thioredoxin reductase and selenoprotein K genes and selenium status modulate risk of prostate cancer. PLoS One. 2012;7(11):e48709. (PubMed)
64. Penney KL, Schumacher FR, Li H, et al. A large prospective study of SEP15 genetic variation, interaction with plasma selenium levels, and prostate cancer risk and survival. Cancer Prev Res (Phila). 2010;3(5):604-610. (PubMed)
65. Yu SY, Zhu YJ, Li WG. Protective role of selenium against hepatitis B virus and primary liver cancer in Qidong. Biol Trace Elem Res. 1997;56(1):117-124. (PubMed)
66. Vinceti M, Crespi CM, Malagoli C, Del Giovane C, Krogh V. Friend or foe? The current epidemiologic evidence on selenium and human cancer risk. J Environ Sci Health C Environ Carcinog Ecotoxicol Rev. 2013;31(4):305-341. (PubMed)
67. Clark LC, Combs GF, Jr., Turnbull BW, et al. Effects of selenium supplementation for cancer prevention in patients with carcinoma of the skin. A randomized controlled trial. Nutritional Prevention of Cancer Study Group. JAMA. 1996;276(24):1957-1963. (PubMed)
68. Reid ME, Duffield-Lillico AJ, Slate E, et al. The nutritional prevention of cancer: 400 μg per day selenium treatment. Nutr Cancer. 2008;60(2):155-163. (PubMed)
69. National Cancer Institute. Review of Prostate Cancer Prevention Study Shows No Benefit for Use of Selenium and Vitamin E Supplements. [Web page]. Available at: http://www.cancer.gov/newscenter/pressreleases/SELECTresults2008. Accessed 10/28/08.
70. Hatfield DL, Gladyshev VN. The Outcome of Selenium and Vitamin E Cancer Prevention Trial (SELECT) reveals the need for better understanding of selenium biology. Mol Interv. 2009;9(1):18-21. (PubMed)
71. Klein EA, Thompson IM, Jr., Tangen CM, et al. Vitamin E and the risk of prostate cancer: the Selenium and Vitamin E Cancer Prevention Trial (SELECT). JAMA. 2011;306(14):1549-1556. (PubMed)
72. Lippman SM, Klein EA, Goodman PJ, et al. Effect of selenium and vitamin E on risk of prostate cancer and other cancers: the Selenium and Vitamin E Cancer Prevention Trial (SELECT). JAMA. 2009;301(1):39-51. (PubMed)
73. Flores-Mateo G, Carrillo-Santisteve P, Elosua R, et al. Antioxidant enzyme activity and coronary heart disease: meta-analyses of observational studies. Am J Epidemiol. 2009;170(2):135-147. (PubMed)
74. Bleys J, Navas-Acien A, Guallar E. Serum selenium levels and all-cause, cancer, and cardiovascular mortality among US adults. Arch Intern Med. 2008;168(4):404-410. (PubMed)
75. Eaton CB, Abdul Baki AR, Waring ME, Roberts MB, Lu B. The association of low selenium and renal insufficiency with coronary heart disease and all-cause mortality: NHANES III follow-up study. Atherosclerosis. 2010;212(2):689-694. (PubMed)
76. Laclaustra M, Navas-Acien A, Stranges S, Ordovas JM, Guallar E. Serum selenium concentrations and hypertension in the US Population. Circ Cardiovasc Qual Outcomes. 2009;2(4):369-376. (PubMed)
77. Kuruppu D, Hendrie HC, Yang L, Gao S. Selenium levels and hypertension: a systematic review of the literature. Public Health Nutr. 2014;17(6):1342-1352. (PubMed)
78. Laclaustra M, Stranges S, Navas-Acien A, Ordovas JM, Guallar E. Serum selenium and serum lipids in US adults: National Health and Nutrition Examination Survey (NHANES) 2003-2004. Atherosclerosis. 2010;210(2):643-648. (PubMed)
79. Stranges S, Laclaustra M, Ji C, et al. Higher selenium status is associated with adverse blood lipid profile in British adults. J Nutr. 2010;140(1):81-87. (PubMed)
80. Rees K, Hartley L, Day C, Flowers N, Clarke A, Stranges S. Selenium supplementation for the primary prevention of cardiovascular disease. Cochrane Database Syst Rev. 2013;1:CD009671. (PubMed)
81. McKenzie RC, Beckett GJ, Arthur JR. Effects of selenium on immunity and aging. In: Hatfield DL, Berry MJ, Gladyshev VN, eds. Selenium: Its Molecular Biology and Role in Human Health. 2nd ed. New York: Springer; 2006:311-323.
82. Huang Z, Rose AH, Hoffmann PR. The role of selenium in inflammation and immunity: from molecular mechanisms to therapeutic opportunities. Antioxid Redox Signal. 2012;16(7):705-743. (PubMed)
83. Mattmiller SA, Carlson BA, Sordillo LM. Regulation of inflammation by selenium and selenoproteins: impact on eicosanoid biosynthesis. J Nutr Sci. 2013;2:e28. (PubMed)
84. Norton RL, Hoffmann PR. Selenium and asthma. Mol Aspects Med. 2012;33(1):98-106. (PubMed)
85. Speckmann B, Steinbrenner H. Selenium and selenoproteins in inflammatory bowel diseases and experimental colitis. Inflamm Bowel Dis. 2014;20(6):1110-1119. (PubMed)
86. Drain PK, Kupka R, Mugusi F, Fawzi WW. Micronutrients in HIV-positive persons receiving highly active antiretroviral therapy. Am J Clin Nutr. 2007;85(2):333-345. (PubMed)
87. Stone CA, Kawai K, Kupka R, Fawzi WW. Role of selenium in HIV infection. Nutr Rev. 2010;68(11):671-681. (PubMed)
88. de Menezes Barbosa EG, Junior FB, Machado AA, Navarro AM. A longer time of exposure to antiretroviral therapy improves selenium levels. Clin Nutr. 2015;34(2):248-251. (PubMed)
89. Baum MK, Miguez-Burbano MJ, Campa A, Shor-Posner G. Selenium and interleukins in persons infected with human immunodeficiency virus type 1. J Infect Dis. 2000;182 Suppl 1:S69-73. (PubMed)
90. Kalantari P, Narayan V, Natarajan SK, et al. Thioredoxin reductase-1 negatively regulates HIV-1 transactivating protein Tat-dependent transcription in human macrophages. J Biol Chem. 2008;283(48):33183-33190. (PubMed)
91. Burbano X, Miguez-Burbano MJ, McCollister K, et al. Impact of a selenium chemoprevention clinical trial on hospital admissions of HIV-infected participants. HIV Clin Trials. 2002;3(6):483-491. (PubMed)
92. Hurwitz BE, Klaus JR, Llabre MM, et al. Suppression of human immunodeficiency virus type 1 viral load with selenium supplementation: a randomized controlled trial. Arch Intern Med. 2007;167(2):148-154. (PubMed)
93. Kupka R, Mugusi F, Aboud S, Hertzmark E, Spiegelman D, Fawzi WW. Effect of selenium supplements on hemoglobin concentration and morbidity among HIV-1-infected Tanzanian women. Clin Infect Dis. 2009;48(10):1475-1478. (PubMed)
94. Kupka R, Mugusi F, Aboud S, et al. Randomized, double-blind, placebo-controlled trial of selenium supplements among HIV-infected pregnant women in Tanzania: effects on maternal and child outcomes. Am J Clin Nutr. 2008;87(6):1802-1808. (PubMed)
95. Baum MK, Campa A, Lai S, et al. Effect of micronutrient supplementation on disease progression in asymptomatic, antiretroviral-naive, HIV-infected adults in Botswana: a randomized clinical trial. JAMA. 2013;310(20):2154-2163. (PubMed)
96. Bone RC, Balk RA, Cerra FB, et al. Definitions for sepsis and organ failure and guidelines for the use of innovative therapies in sepsis. The ACCP/SCCM Consensus Conference Committee. American College of Chest Physicians/Society of Critical Care Medicine. Chest. 1992;101(6):1644-1655. (PubMed)
97. Mann EA, Baun MM, Meininger JC, Wade CE. Comparison of mortality associated with sepsis in the burn, trauma, and general intensive care unit patient: a systematic review of the literature. Shock. 2012;37(1):4-16. (PubMed)
98. Alhazzani W, Jacobi J, Sindi A, et al. The effect of selenium therapy on mortality in patients with sepsis syndrome: a systematic review and meta-analysis of randomized controlled trials. Crit Care Med. 2013;41(6):1555-1564. (PubMed)
99. Huang TS, Shyu YC, Chen HY, et al. Effect of parenteral selenium supplementation in critically ill patients: a systematic review and meta-analysis. PLoS One. 2013;8(1):e54431. (PubMed)
100. van Zuuren EJ, Albusta AY, Fedorowicz Z, Carter B, Pijl H. Selenium Supplementation for Hashimoto's Thyroiditis: Summary of a Cochrane Systematic Review. Eur Thyroid J. 2014;3(1):25-31. (PubMed)
101. Karanikas G, Schuetz M, Kontur S, et al. No immunological benefit of selenium in consecutive patients with autoimmune thyroiditis. Thyroid. 2008;18(1):7-12. (PubMed)
102. Krysiak R, Okopien B. The effect of levothyroxine and selenomethionine on lymphocyte and monocyte cytokine release in women with Hashimoto's thyroiditis. J Clin Endocrinol Metab. 2011;96(7):2206-2215. (PubMed)
103. Negro R, Greco G, Mangieri T, Pezzarossa A, Dazzi D, Hassan H. The influence of selenium supplementation on postpartum thyroid status in pregnant women with thyroid peroxidase autoantibodies. J Clin Endocrinol Metab. 2007;92(4):1263-1268. (PubMed)
104. Turker O, Kumanlioglu K, Karapolat I, Dogan I. Selenium treatment in autoimmune thyroiditis: 9-month follow-up with variable doses. J Endocrinol. 2006;190(1):151-156. (PubMed)
105. Marcocci C, Kahaly GJ, Krassas GE, et al. Selenium and the course of mild Graves' orbitopathy. N Engl J Med. 2011;364(20):1920-1931. (PubMed)
106. Watt T, Cramon P, Bjorner JB, et al. Selenium supplementation for patients with Graves' hyperthyroidism (the GRASS trial): study protocol for a randomized controlled trial. Trials. 2013;14:119. (PubMed)
107. Winther KH, Watt T, Bjorner JB, et al. The chronic autoimmune thyroiditis quality of life selenium trial (CATALYST): study protocol for a randomized controlled trial. Trials. 2014;15:115. (PubMed)
108. Peplow D, Edmonds R. Health risks associated with contamination of groundwater by abandoned mines near Twisp in Okanogan County, Washington, USA. Environ Geochem Health. 2004;26(1):69-79. (PubMed)
109. Chang JC, Gutenmann WH, Reid CM, Lisk DJ. Selenium content of Brazil nuts from two geographic locations in Brazil. Chemosphere. 1995;30(4):801-802. (PubMed)
110. Rayman MP, Infante HG, Sargent M. Food-chain selenium and human health: spotlight on speciation. Br J Nutr. 2008;100(2):238-253. (PubMed)
111. Reyes LH, Encinar JR, Marchante-Gayon JM, Alonso JI, Sanz-Medel A. Selenium bioaccessibility assessment in selenized yeast after "in vitro" gastrointestinal digestion using two-dimensional chromatography and mass spectrometry. J Chromatogr A. 2006;1110(1-2):108-116. (PubMed)
112. Bermingham EN, Hesketh JE, Sinclair BR, Koolaard JP, Roy NC. Selenium-Enriched Foods Are More Effective at Increasing Glutathione Peroxidase (GPx) Activity Compared with Selenomethionine: A Meta-Analysis. Nutrients. 2014;6(10):4002-4031. (PubMed)
113. Giacosa A, Faliva MA, Perna S, Minoia C, Ronchi A, Rondanelli M. Selenium fortification of an Italian rice cultivar via foliar fertilization with sodium selenate and its effects on human serum selenium levels and on erythrocyte glutathione peroxidase activity. Nutrients. 2014;6(3):1251-1261. (PubMed)
114. Bleys J, Navas-Acien A, Guallar E. Serum selenium and diabetes in U.S. adults. Diabetes Care. 2007;30(4):829-834. (PubMed)
115. Laclaustra M, Navas-Acien A, Stranges S, Ordovas JM, Guallar E. Serum selenium concentrations and diabetes in U.S. adults: National Health and Nutrition Examination Survey (NHANES) 2003-2004. Environ Health Perspect. 2009;117(9):1409-1413. (PubMed)
116. Stranges S, Marshall JR, Natarajan R, et al. Effects of Long-Term Selenium Supplementation on the Incidence of Type 2 Diabetes: A Randomized Trial. Ann Intern Med. 2007;147(4):217-223. (PubMed)
117. Schoenmakers E, Agostini M, Mitchell C, et al. Mutations in the selenocysteine insertion sequence-binding protein 2 gene lead to a multisystem selenoprotein deficiency disorder in humans. J Clin Invest. 2010;120(12):4220-4235. (PubMed)
118. Steinbrenner H. Interference of selenium and selenoproteins with the insulin-regulated carbohydrate and lipid metabolism. Free Radic Biol Med. 2013;65:1538-1547. (PubMed)
119. Kaur P, Rizk NM, Ibrahim S, et al. iTRAQ-based quantitative protein expression profiling and MRM verification of markers in type 2 diabetes. J Proteome Res. 2012;11(11):5527-5539. (PubMed)
120. Yang SJ, Hwang SY, Choi HY, et al. Serum selenoprotein P levels in patients with type 2 diabetes and prediabetes: implications for insulin resistance, inflammation, and atherosclerosis. J Clin Endocrinol Metab. 2011;96(8):E1325-1329. (PubMed)
121. Azrak RG, Cao S, Pendyala L, et al. Efficacy of increasing the therapeutic index of irinotecan, plasma and tissue selenium concentrations is methylselenocysteine dose dependent. Biochem Pharmacol. 2007;73(9):1280-1287. (PubMed)
122. Graf WD, Oleinik OE, Glauser TA, Maertens P, Eder DN, Pippenger CE. Altered antioxidant enzyme activities in children with a serious adverse experience related to valproic acid therapy. Neuropediatrics. 1998;29(4):195-201. (PubMed)
123. Vernie LN, de Goeij JJ, Zegers C, de Vries M, Baldew GS, McVie JG. Cisplatin-induced changes of selenium levels and glutathione peroxidase activities in blood of testis tumor patients. Cancer Lett. 1988;40(1):83-91. (PubMed)
124. Flodin NW. Micronutrient supplements: toxicity and drug interactions. Prog Food Nutr Sci. 1990;14(4):277-331. (PubMed)
125. Brown BG, Zhao XQ, Chait A, et al. Simvastatin and niacin, antioxidant vitamins, or the combination for the prevention of coronary disease. N Engl J Med. 2001;345(22):1583-1592. (PubMed)
126. Tousoulis D, Antoniades C, Stefanadis C. Statins and antioxidant vitamins: should co-administration be avoided? J Am Coll Cardiol. 2006;47(6):1237; author reply 1237-1238. (PubMed)
127. Hurst R, Armah CN, Dainty JR, et al. Establishing optimal selenium status: results of a randomized, double-blind, placebo-controlled trial. Am J Clin Nutr. 2010;91(4):923-931. (PubMed)
128. Burk RF, Norsworthy BK, Hill KE, Motley AK, Byrne DW. Effects of chemical form of selenium on plasma biomarkers in a high-dose human supplementation trial. Cancer Epidemiol Biomarkers Prev. 2006;15(4):804-810. (PubMed)
129. Algotar AM, Stratton MS, Ahmann FR, et al. Phase 3 clinical trial investigating the effect of selenium supplementation in men at high-risk for prostate cancer. Prostate. 2013;73(3):328-335. (PubMed)
130. Chiang EC, Shen S, Kengeri SS, et al. Defining the Optimal Selenium Dose for Prostate Cancer Risk Reduction: Insights from the U-Shaped Relationship between Selenium Status, DNA Damage, and Apoptosis. Dose Response. 2009;8(3):285-300. (PubMed)
131. Ravn-Haren G, Krath BN, Overvad K, et al. Effect of long-term selenium yeast intervention on activity and gene expression of antioxidant and xenobiotic metabolising enzymes in healthy elderly volunteers from the Danish Prevention of Cancer by Intervention by Selenium (PRECISE) pilot study. Br J Nutr. 2008;99(6):1190-1198. (PubMed)






