Resumen
- La riboflavina es el precursor de las coenzimas, flavín adenín dinucleótido (FAD) y flavín mononucleótido (FMN). Estas actúan como transportadores de electrones en un cierto número de reacciones de oxidación-reducción (redox) involucradas en la producción de energía y en numerosas rutas metabólicas. (Más información)
- La deficiencia de riboflavina puede afectar múltiples rutas en el metabolismo de la vitamina B6, folato, niacina, y del hierro. (Más información)
- La deficiencia de riboflavina ha sido ligada a la preeclampsia en mujeres embarazadas. Esta condición puede progresar a eclampsia y causar hemorragias severas y la muerte. El riesgo de preeclampsia ha sido recientemente asociado con la presencia de una variante genética (C677T) en el gene de la metilentetrahidrofolato reductasa (MTHFR). Este gene codifica para la enzima MTHFR, la cual es dependiente de FAD. (Más información)
- El estrés oxidativo puede causar opacificación del cristalino del ojo, conduciendo a las cataratas en individuos mayores. Mientras los resultados de algunos estudios basados en la observación son prometedores, estudios de intervención son necesarios para evaluar el beneficio potencial de la riboflavina en la prevención de cataratas. (Más información)
- La riboflavina (en la forma de FAD) es requerida como cofactor para la enzima clave que metaboliza el folato, MTHFR. Un estatus bajo del estatus de la riboflavina puede interferir con el metabolismo del folato, particularmente en individuos homocigotos para la variante del gen MTHFR C677T; estos individuos exhiben un riesgo mayor de enfermedades cardiovasculares (ECV). Evidencia emergente proveniente de ensayos de intervención apoya un papel protector de la riboflavina contra la hipertensión en individuos con el genotipo MTHFR 677TT. (Más información)
- Se ha evaluado a la riboflavina como un agente profiláctico en estudios con niños y adultos que padecen de migraña. La suplementación con riboflavina ha mostrado disminuir la frecuencia y severidad de los ataques de dolor de cabeza en adultos pero no en niños. (Más información)
- Reportes de caso han mostrado que los pacientes con trastornos autosómicos recesivos del metabolismo de riboflavina podrían beneficiarse de la suplementación con riboflavina. (Más información)
- Se ha evaluado a la riboflavina como una terapia adyuvante potencial en el cáncer y ciertos desordenes oculares. (Más información)

La riboflavina es una vitamina B hidrosoluble, también conocida como vitamina B2. En el cuerpo, la riboflavina es principalmente encontrada como un componente integral de las coenzimas, flavín adenín dinucleótido (FAD) y flavín mononucleótido (FMN) (1). Las coenzimas derivadas de la riboflavina se denominan flavocoenzimas, y enzimas que usan una flavocoenzima son llamadas flavoproteínas (2).
Función
Reacciones de oxidación-reducción (redox)
Los organismos vivos derivan la mayoría de su energía de las reacciones redox, las cuales son procesos que involucran la transferencia de electrones. Las flavocoenzimas participan en las reacciones redox en numerosas rutas metabólicas (3). Son críticas para el metabolismo de los carbohidratos, lípidos, y proteínas. El FAD es parte de la cadena de transporte (respiratoria) de electrones, la cual es fundamental para la producción de energía. En conjunción con el citocromo P-450, las flavocoenzimas también participan en el metabolismo de drogas y toxinas (4).
Funciones antioxidantes
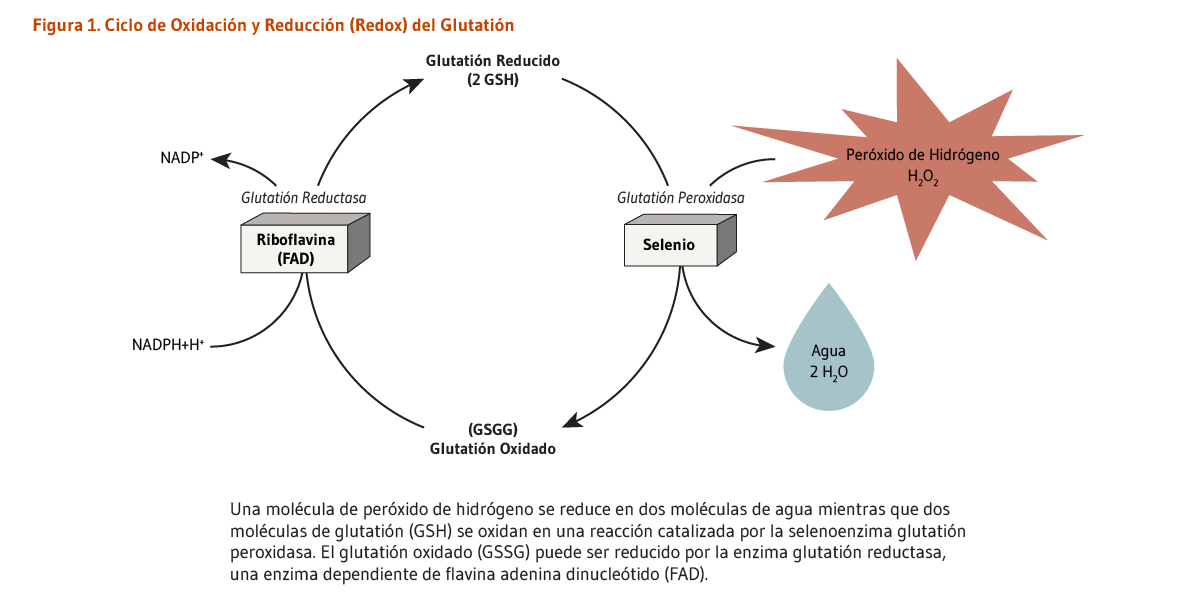
La glutatión reductasa es una enzima dependiente de FAD que participa en el ciclo redox del glutatión. El ciclo redox del glutatión juega un papel fundamental en la protección de organismos de especies reactivas del oxígeno, como los hidroperóxidos. La glutatión reductasa (GR) requiere de FAD para regenerar dos moléculas de glutatión reducido a partir del glutatión oxidado. La deficiencia de riboflavina ha sido asociada con un incremento del estrés oxidativo (4). La medición de la actividad de la GR en los glóbulos rojos es comúnmente usada para evaluar el estatus nutricional de la riboflavina (5). El ensayo del coeficiente de activación de la glutatión reductasa eritrocitaria (EGRAC) evalúa el estatus de la riboflavina al medir la actividad de la GR antes y después de la reactivación in vitro con su grupo prostético FAD; el EGRAC es calculado como la proporción de FAD-estimulado con respecto a la actividad enzimática no estimulada e indica el grado de saturación con riboflavina del tejido. De esta manera el EGRAC es una medida fundamental del estatus de riboflavina y ha mostrado ser efectiva en reflejar el estatus del biomarcador de una deficiencia severa a un estatus normal (6).
Las glutatión peroxidasas, enzimas que contienen selenio, necesitan de dos moléculas de glutatión reducido para degradar hidroperóxidos. Las GPx están involucradas en el ciclo de oxidación-reducción (redox) del glutatión (Figura 1).
La xantina oxidasa, otra enzima dependiente de FAD, cataliza la oxidación de hipoxantina y xantina a ácido úrico. El ácido úrico es uno de los antioxidantes hidrosolubles más efectivos en la sangre. Por lo tanto, la deficiencia de riboflavina puede derivar en una actividad disminuida de la xantina oxidasa, reduciendo los niveles sanguíneos de ácido úrico (7).
[Figura 1 - Clic para Agrandar]
Interacción con nutrientes
Vitaminas del complejo B
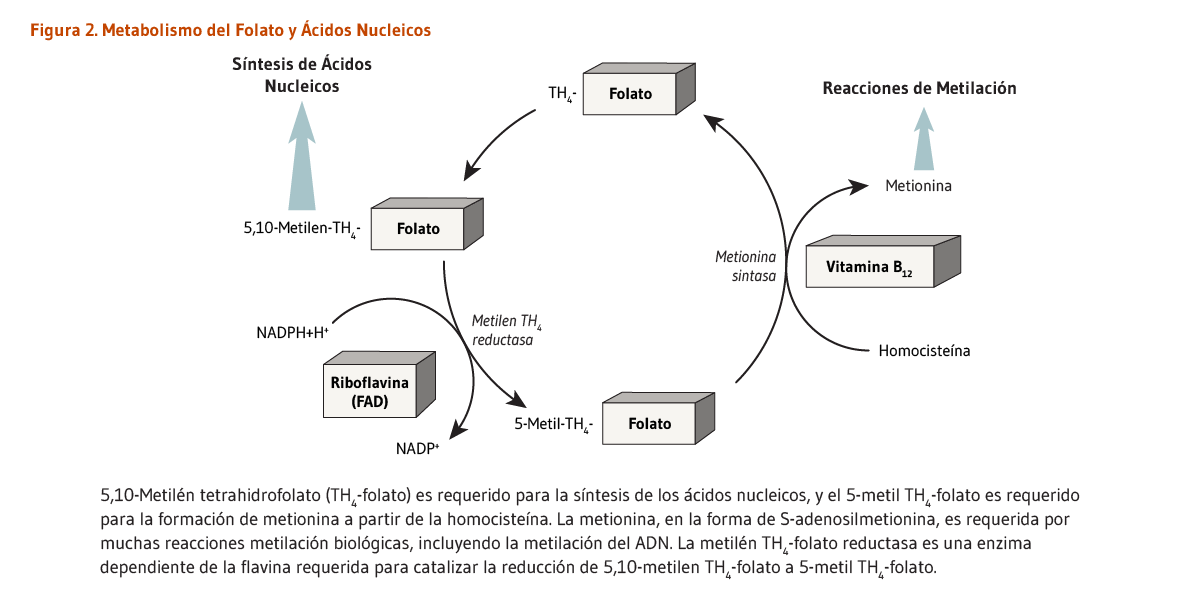
Las flavoproteínas están involucradas en el metabolismo de varias otras vitaminas: (vitamina B6, niacina, y folato). Por lo tanto, una deficiencia severa de riboflavina podría afectar muchos sistemas de enzimas. La conversión de la mayoría de vitamina B6 naturalmente disponible a su forma de coenzima, piridoxal 5’-fosfato (PLP), requiere de la enzima dependiente de FMN, piridoxina 5’-fosfato oxidasa (PPO) (8). Por lo menos dos estudios en adultos mayores han documentado interacciones significantes entre los indicadores de vitamina B6 y el estatus nutricional de la riboflavina (9, 10). La síntesis de coenzimas que contienen niacina, NAD y NADP, a partir del aminoácido triptófano, requiere de la enzima dependiente de FAD, quinurenina mono-oxigenasa. Una deficiencia severa de riboflavina puede disminuir la conversión de triptófano a NAD y NADP, incrementando el riesgo de una deficiencia de niacina (3). La 5,10-metilentetrahidrofolato reductasa (MTHFR) es una enzima dependiente de FAD que juega un papel importante en el mantenimiento de una coenzima folato específica, requerida para formar metionina a partir de homocisteína (Figura 2). Junto con otras vitaminas del complejo B, se ha asociado a las ingestas elevadas de riboflavina con niveles disminuidos de homocisteína en el plasma (11). Niveles incrementados de riboflavina en el plasma han sido también asociados con niveles disminuidos de homocisteína del plasma, principalmente en individuos homocigotos para el polimorfismo C677T en el gen de la MTHFR y en individuos con una baja ingesta de folato (12). Tales resultados muestran que el riesgo de enfermedades crónicas podría ser influenciado por interacciones complejas entre factores genéticos y dietarios (véase Enfermedades cardiovasculares y Cáncer).
[Figura 2 - Clic para Agrandar]
Hierro
La deficiencia de riboflavina altera el metabolismo del hierro. Aunque el mecanismo no está aún claro, investigación en animales sugiere que la deficiencia de riboflavina podría perjudicar la absorción del hierro, incrementar la pérdida intestinal de hierro y/o afectar la utilización del hierro para la síntesis de hemoglobina (Hb) (13). En humanos, se ha encontrado que al mejorar el estatus nutricional de la riboflavina se aumentan los niveles de Hb circulante (14). La corrección de una deficiencia de riboflavina en individuos que son deficientes de ambos riboflavina y hierro mejora la respuesta de individuos con anemia ferropénica a la terapia con hierro (15). La anemia durante el embarazo, un problema de salud pública alrededor del mundo, es responsable por una considerable morbilidad y mortalidad perinatal (16, 17). El manejo de la anemia materna incluye la suplementación con hierro solo o hierro en combinación con ácido fólico (18), y se ha considerado que la suplementación con riboflavina podría mejorar la suplementación con hierro-ácido fólico. Ensayos de intervención, aleatorios, doble ciegos conducidos en mujeres embarazadas con anemia en el Sureste Asiático mostraron que una combinación de ácido fólico, hierro, vitamina A, y riboflavina mejoró los niveles de Hb y disminuyó la prevalencia de anemia en comparación a la suplementación sola con hierro-ácido fólico (19, 20).
Deficiencia
Ariboflavonosis es el término médico para una deficiencia clínica de riboflavina. Una deficiencia de riboflavina es raramente encontrada de manera aislada; ocurre con frecuencia en combinación con deficiencia de otras vitaminas hidrosolubles. Los síntomas de una deficiencia de riboflavina incluyen dolor de garganta, enrojecimiento e hinchazón de la mucosa de la boca y garganta, grietas o llagas en el exterior de los labios (quelosis) y en las comisuras de la boca (estomatitis angular), inflamación y enrojecimiento de la lengua (lengua magenta), y una inflamación cutánea húmeda y escamosa (dermatitis seborreica). Otros síntomas podrían involucrar la formación de vasos sanguíneos en la capa transparente del ojo (vascularización de la córnea) y conteo disminuido de glóbulos rojos en el cual los glóbulos rojos existentes contienen niveles normales de hemoglobina y son de tamaño normal (anemia normocítica normocrómica) (1, 3). Una deficiencia severa de riboflavina podría resultar en la conversión disminuida de vitamina B6 a su forma de coenzima (PLP) y una conversión disminuida de triptófano a niacina (véase Interacciones con nutrientes).
La preeclampsia es definida como la presencia de presión sanguínea elevada, proteínas en la orina y edema (hinchazón significativa) durante el embarazo. Cerca del 5% de las mujeres con preeclampsia progresan a eclampsia, una causa importante de muerte materna y fetal. La eclampsia es caracterizada por convulsiones, además de presión sanguínea alta y un riesgo aumentado de hemorragia (sangramiento severo) (21). Un estudio en 154 mujeres embarazadas en riesgo incrementado de preeclampsia encontró que aquellas que estaban deficientes de riboflavina eran 4.7 veces más propensas a desarrollar preeclampsia que aquellas que tenían un estatus nutricional adecuado de riboflavina (22). La causa de la preeclampsia-eclampsia es desconocida. Niveles intracelulares de flavocoenzimas disminuidos podrían causar disfunción mitocondrial, aumento del estrés oxidativo e interferencia con la liberación de óxido nítrico y, por lo tanto, con la dilatación de vasos sanguíneos — todos estos cambios han sido asociados con la preeclampsia (22).
Un reciente meta-análisis de 51 estudios encontró que el polimorfismo C677T del gen de la metilentetrahidrofolato reductasa (MTHFR) estaba asociado con la preeclamsia en las poblaciones caucásicas y del Este de Asia (23). Sin embargo un meta-análisis temprano reportó que el genotipo 677TT MTHFR acarreaba un riesgo significantemente mayor de preeclampsia entre mujeres asiáticas únicamente, mientras que en mujeres caucásicas el incremento del riesgo no fue significante (24). Tal heterogeneidad entre estudios podría sugerir que el efecto del genotipo 677TT MTHFR podría ser modulado por la riboflavina y otros factores dietarios relevantes que podrían variar considerablemente entre las diferentes poblaciones. La reducción en la actividad de la flavoproteína de la MTHFR observada en sujetos con la variante genética C677T conduce a un incremento leve en las concentraciones de homocisteína del plasma; los niveles incrementados de homocisteína han sido asociados con la preeclampsia (25). Un ensayo pequeño doble ciego, aleatorio, controlado con placebo en 450 mujeres embarazadas en alto riesgo de preeclampsia encontró que la suplementación con 15 mg de riboflavina diariamente no impidió la condición (26). Sin embargo, estudios son necesarios para evaluar el beneficio potencial de la suplementación con riboflavina en la reducción de complicaciones perinatales específicamente en mujeres preeclámpticas con el genotipo C677T.
Factores de riesgo para la deficiencia de riboflavina
Los alcohólicos están en un riesgo incrementado de deficiencia de riboflavina debido a una ingesta disminuida, una absorción disminuida y a la utilización deteriorada de la riboflavina. Curiosamente, los niveles elevados de homocisteína asociados con la deficiencia de riboflavina rápidamente declinaron durante la abstinencia de alcohol (27). Adicionalmente, las personas anoréxicas rara vez consumen riboflavina adecuada, e individuos con intolerancia a la lactosa podrían no consumir leche u otros productos lácteos que son buenas fuentes de riboflavina. La conversión de riboflavina a FAD y FMN es alterada en el hipotiroidismo e insuficiencia adrenal (3, 4). Además, las personas que son muy activas físicamente (atletas, trabajadores) pudiesen tener un requerimiento de riboflavina levemente incrementado. Sin embargo, generalmente no se ha encontrado que la suplementación con riboflavina incremente el desempeño o la tolerancia al ejercicio (28).
Aunque la deficiencia clínica de riboflavina (es decir, incluyendo signos tales como la estomatitis angular, queilosis, y glositis) es rara en el mundo desarrollado, existe evidencia que sugiere que el estatus subóptimo de la riboflavina (determinado por el biomarcador funcional EGRAC) podría ser un problema generalizado que afecta a muchas poblaciones de otro modo sanas dentro del mundo desarrollado. Por ejemplo, se reportó que una proporción alta de la población británica adulta poseía un estatus de riboflavina pobre como se determinó de los datos de la encuesta nacional usando EGRAC (29). Las ingestas dietarías sin embargo fueron generalmente favorables al ser comparadas con los valores recomendados, excepto en mujeres jóvenes que tuvieron ingestas bajas. La gran proporción de la población británica con valores anormales de EGRAC, a pesar de ingestas dietarías aparentemente adecuadas, requiere de investigación adicional.
La IDR
La ingesta diaria recomendada (IDR) para la riboflavina, revisada en 1998, está basada en la prevención de su deficiencia (Tabla 1). En humanos los signos clínicos de la deficiencia aparecen a ingestas de menos de 0.5-0.6 miligramos (mg)/día, y la excreción urinaria de riboflavina es vista a niveles de ingesta de aproximadamente 1 mg/día (1).
| Etapa de la Vida | Edad | Machos (mg/día) | Hembras (mg/día) |
|---|---|---|---|
| Infantes | 0-6 meses | 0.3 (IA) | 0.3 (IA) |
| Infantes | 7-12 meses | 0.4 (IA) | 0.4 (IA) |
| Niños | 1-3 años | 0.5 | 0.5 |
| Niños | 4-8 años | 0.6 | 0.6 |
| Niños | 9-13 años | 0.9 | 0.9 |
| Adolescentes | 14-18 años | 1.3 | 1.0 |
| Adultos | 19 años y más | 1.3 | 1.1 |
| Embarazo | Todas las edades | - | 1.4 |
| Período de lactancia | Todas las edades | - | 1.6 |
Prevención de Enfermedades
Cataratas
Las cataratas seniles son la causa principal de discapacidad visual en los EE.UU. y otros países desarrollados. La investigación se ha enfocado en el papel de los antioxidantes nutricionales debido a la evidencia de que el daño oxidativo foto-inducido de las proteínas del cristalino podría conducir al desarrollo de cataratas relacionadas con la edad. Un estudio de caso y control encontró un riesgo significativamente disminuido de cataratas seniles (33 a 51%) en hombres y mujeres en el quintil más alto de la ingesta de riboflavina (mediana de 1.6 a 2.2 mg/día) en comparación a aquellos en el quintil más bajo (mediana de 0.08 mg/día tanto en hombres como en mujeres) (30). Otro estudio de caso y control reportó que los individuos en el quintil más elevado del estatus nutricional de la riboflavina, medido por la actividad de la glutatión reductasa de los glóbulos rojos, tenían aproximadamente la mitad de la incidencia de cataratas seniles que aquellos en el quintil más bajo del estatus de riboflavina, aunque estos resultados no fueron estadísticamente significativos (31). Un estudio de corte transversal de 2,900 hombres y mujeres australianos, de 49 años de edad y mayores, encontró que aquellos en el quintil más alto de la ingesta de riboflavina eran un 50% menos propensos a tener cataratas que aquellos en el quintil más bajo (32). Un estudio prospectivo de más de 50,000 mujeres no observo alguna diferencia entre las tasas de extracción de cataratas entre mujeres en el quintil más alto de la ingesta de riboflavina (mediana de 1.5 mg/día) y mujeres en el quintil más bajo (mediana de 1.2 mg/día) (33). Sin embargo, el rango entre el quintil más alto y el más bajo era pequeño, y los niveles de ingesta medianos para ambos quintiles estaban por sobre la actual IDR para la riboflavina. Un estudio en 480 mujeres encontró que ingestas recomendada más altas de riboflavina se asociaban inversamente con cambios en la opacificación de los cristalinos en un período de cinco años (34). Aunque estos estudios basados en la observación proporcionan apoyo al papel de la riboflavina en la prevención de las cataratas, es necesario realizar ensayos de intervención aleatorios, controlados con placebo que incluyan un biomacador de respuesta (como la EGRAC) para confirmar esta relación.
Enfermedades cardiovasculares
Por muchos años, los niveles elevados de homocisteína en el plasma han sido considerados como un factor de riesgo para las enfermedades cardiovasculares (ECV), aunque esto recientemente se ha convertido de alguna manera en algo controversial (35). La homocisteína del plasma responde a los efectos reductores de las intervenciones con folato y vitaminas B relacionadas metabólicamente, incluyendo la riboflavina. La riboflavina actúa como un cofactor para la MTHFR y es por ello necesaria para generar 5-metiltetrahidrofolato requerido en la remetilación de la homocisteína a metionina (véase Figura 2 arriba). Estas vitaminas B, sin embargo, podrían tener papeles en la prevención de ECV que son independientes de sus efectos sobre la homocisteína.
Estudios genéticos proporcionan evidencia convincente para apoyar un vínculo entre el estatus sub-óptimo de vitamina B y el riesgo de ECV. Un meta-análisis de tales estudios mostró que los individuos que son homocigotos para la variante del gen MTHFR C677T tuvieron un riesgo significantemente mayor de ECV (por un 14% a 21%) en comparación a aquellos sin este polimorfismo, pero hubo una gran cantidad de variación geográfica en el incremento del riesgo de ECV (36), sugiriendo fuertemente que los factores dietarios pueden modular el riesgo de la enfermedad relacionado a este factor genético.
La acumulación de pruebas vincula este polimorfismo común del folato con la hipertensión (definida como una presión sanguínea de 140/90 mm Hg o mayor), un factor de riesgo principal para las ECV, particularmente los accidentes cerebrovasculares. La evidencia emergente de que la presión sanguínea en pacientes homocigotos para este polimorfismo es altamente sensible a dosis bajas de riboflavina (véase Tratamiento de Enfermedades) aumenta la posibilidad de que el mejoramiento del estatus de la riboflavina tendrá un papel importante en la prevención de la hipertensión. Esto en turno podría potencialmente reducir el riesgo de accidentes cerebrovasculares específicamente en individuos con el genotipo relevante. Notablemente, la frecuencia reportada del genotipo MTHFR C677T es de un 10% en todo el mundo, oscilando entre 4%-26% en Europa, 20% en el norte de China, y tan alto como un 32% en México (37).
Cáncer
La flavoproteína, metilentetrahidrofolato reductasa (MTHFR), juega un papel fundamental en el metabolismo de la homocisteína mediada por el folato. La MTHFR convierte el 5,10-metilentetrahidrofolato a 5-metiltetrahidrofolato, el cual es necesario para le re-metilación de la homocisteína a metionina (véase Figura 2 arriba). La conversión de la homocisteína a metionina es de importancia para la desintoxicación de la homocisteína y para la producción de S-adenosilmetionina (SAM), el donante de metilo para la metilación del ADN y las histonas. La deficiencia de folato y elevadas concentraciones de homocisteína podrían incrementar el riesgo de cáncer (véase el artículo en Folato). Los cambios aberrantes en la metilación, son también conocidos por alterar la estructura y función del ADN y las histonas durante el desarrollo del cáncer (38). Ya que la MTHFR controla la desintoxicación de la homocisteína y el suministro de grupos de metilo para la síntesis de SAM, una reducción en su actividad puede afectar el metabolismo de la homocisteína y alterar los procesos de metilación celular. La sustitución de una citosina por una timina en la posición 677 (c.677C>T) en el gen de la MTHFR es un polimorfismo que afecta la unión de FAD y conduce a una mayor propensión de la MTHFR a perder su coenzima flavina (39). Individuos homocigotos para esta mutación (es decir, genotipo MTHFR 677TT) exhiben una actividad de la MTHFR reducida, y evidencia demuestra que tales individuos están un riesgo incrementado de cáncer en varios sitios (40-42); sin embargo, la naturaleza de la asociación entre este polimorfismo común y el riesgo de cáncer permanece incierto.
Como se mencionó previamente (véase Vitaminas del complejo B), la ingesta de riboflavina es un determinante de la concentración de homocisteína. Esto sugiere que el estatus de la riboflavina puede influenciar la actividad de la MTHFR y el metabolismo del folato, afectando potencialmente así el riesgo de cáncer (42). En un estudio aleatorio, doble ciego, controlado con placebo, a 93 sujetos con pólipos colorrectales y 86 sujetos saludable se les dio o un placebo, ácido fólico (400 o 1,200 mcg/día), o ácido fólico (400 mcg/día) más riboflavina (5 mg/día) por 45 días. Estas intervenciones significantemente mejoraron el estatus del folato y la riboflavina en los individuos suplementados con vitaminas en comparación con aquellos que tomaron el placebo. Curiosamente, la riboflavina mejoró el efecto de 400 mcg de ácido fólico en la 5-metil tetrahidrafolato (5-MeTH4) circulante específicamente en los pacientes con pólipos con la variante genética C677T (43). Esto sugiere que la riboflavina pudiese mejorar la respuesta a la suplementación con ácido fólico en individuos con una actividad de la MTHFR reducida. Adicionalmente, un estudio de cohorte prospectivo de 88,045 mujeres postmenopáusicas encontró que la ingesta total (dietaría más suplementaria) de riboflavina estaba inversamente correlacionada con el riesgo de cáncer colorrectal cuando se compararon los cuartiles más altos (>3.97 mg) y los más bajo (<1.80 mg) de la ingesta diaria (44); la ingesta en el grupo de referencia estaba muy por arriba de la IDR de 1.1 mg/día. Los sujetos en este estudio no fueron pre-cribados para identificar aquellos con el genotipo C677T, y la asociación entre el polimorfismo C677T y el cáncer colorrectal no es claro, con reportes sugiriendo una reducción en el riesgo con el alelo T (45).
Asociaciones entre la ingesta de riboflavina y el riesgo de cáncer han sido evaluadas en otros tipos de cáncer. Un estudio de intervención de siete años evaluó el uso de sal fortificada con riboflavina en 22,093 individuos en un alto riesgo de cáncer de esófago en China. El estatus de la riboflavina y la patología esofágica (porcentaje de tejidos normales, displásicos y cancerosos) mejoró en el grupo de intervención en comparación al grupo de control, pero la menor incidencia de cáncer de esófago encontrada en el grupo de intervención no fue estadísticamente significante (46). Adicionalmente, el Estudio de Cohorte Colaborativo de Melbourne, el cual dio seguimiento a 41,514 hombres y mujeres sobre un periodo de 15 años, encontró asociaciones inversas débiles entre la ingesta de riboflavina y el cáncer de pulmón (47) y el cáncer de seno (48) y no asociación alguna con el cáncer de próstata (49). Sin embargo, un seguimiento de 10 años de un ensayo de intervención en pacientes en alto riesgo de cáncer gástrico (estomago) encontró que la suplementación dietaría con minerales y vitaminas, incluyendo riboflavina (3.2 mg/día) y niacina (40 mg/día), por cinco años falló en disminuir la incidencia o la tasa de mortalidad del cáncer gástrico (50).
Tratamiento de Enfermedades
Migrañas
Cierta evidencia indica que un metabolismo mitocondrial del oxígeno deteriorado en el cerebro podría jugar un papel en la patología de las migrañas. Debido a que la riboflavina es el precursor de dos flavocoenzimas (FAD y FMN) requeridas por las flavoproteínas de la cadena transportadora de electrones mitocondrial, se ha investigado a la riboflavina suplementaria como un tratamiento para la migraña. Un ensayo aleatorio, controlado con placebo examinó el efecto de 400 mg/día de riboflavina al día por tres meses en la prevención de migraña en 54 hombres y mujeres con un historial de migrañas recurrentes (51). La riboflavina fue significativamente mejor que el placebo en la reducción de la frecuencia de los ataques y el número de días con dolores de cabeza, aunque el efecto benéfico fue más pronunciado durante el tercer mes de tratamiento. Otro estudio por los mismos investigadores encontró que el tratamiento tanto con un medicamento β-bloqueador o con riboflavina en dosis elevadas (400 mg/día) por cuatro meses resultó en una mejora clínica, pero cada terapia pareció actuar sobre un mecanismo patológico diferente: los β-bloqueadores en el procesamiento de información cortical anormal y la riboflavina sobre las reservas cerebrales de energía mitocondrial disminuidas (52). Un estudio pequeño en 23 pacientes reportó una reducción en la frecuencia media de ataques de migraña después de la suplementación con 400 mg de riboflavina diariamente por tres meses (53). Sin embargo, un estudio de 3 meses, aleatorio, doble ciego, controlado con placebo que administró una combinación de riboflavina (400 mg/día), magnesio, y matricaria a personas que sufren de migraña reportó ningún beneficio terapéutico más allá del asociado con consumir un placebo que contenga 25 mg/día de riboflavina (54). En comparación con las mediciones basales en este estudio, tanto el grupo que recibió el placebo como el grupo en tratamiento experimentaron ciertos beneficios con respecto al promedio de migrañas, días con migraña, o en el índice de las migrañas (54). Aunque estos resultados son preliminares, los datos procedentes de la mayoría de los estudios hasta la fecha sugieren que la suplementación con riboflavina en adultos podría ser un complemento útil para la terapia farmacológica en la prevención de migrañas.
Dos ensayos aleatorios, doble ciegos, controlados con placebo investigaron el efecto de la suplementación con riboflavina en la frecuencia y severidad de los ataques de dolor de cabeza en niños con migrañas. El primer estudio evaluó la riboflavina en 200 mg/día por 12 semanas en 48 niños de entre 5 a 15 años de edad (55). El segundo estudio fue un ensayo cruzado con la mitad de los 42 niños, de entre 6 a 13 años de edad, recibiendo 50 mg/día de riboflavina por 16 semanas luego placebo (100 mg/día caroteno) por 16 semanas con un periodo de lavado de cuatro semanas de por medio, mientras que a la otra mitad se les dio primero placebo luego riboflavina (56). Ambos estudios no mostraron diferencias en la frecuencia, duración, o intensidad de migrañas entre tratamientos. La efectividad de la suplementación con riboflavina en niños con migraña no es aun establecida, e investigación futura debe primero determinar la dosis más apropiada de riboflavina para esta población.
Trastornos metabólicos
Evidencia creciente indica que pacientes con trastornos autosómicos recesivos causados por enzimas dependientes de FAD defectuosas podrían beneficiarse de la suplementación con riboflavina.
Deficiencia múltiple de acil CoA-deshidrogenasa (MADD)
MADD, también conocida como aciduria glutárica tipo II (o acidemia), es un trastorno del metabolismo de ácidos grasos caracterizado por la acumulación de acil-carnitias de cadena corta, mediana, y larga en varios tejidos. Formas neonatales y de aparición tardía de la MADD muestran un amplio espectro de severidad clínica y presentaciones variables, incluyendo encefalopatía episódica, vómitos periódicos, hepatopatía y rabdomiólisis (57). La MADD es causada por mutaciones en genes que perjudican la actividad de enzimas involucradas en la transferencia de electrones a partir de la acil-Coenzima A (acil-CoA) a la Coenzima Q10 ubiquinona dentro de la mitocondria (Figura 3). ETFA, ETFB, y ETFDH codifican para las dos subunidades de la flavoproteína de transferencia de electrones (ETF-A y -B), y para la ETF deshidrogenasa/ubiquinona oxidorreductasa (ETFDH/ETFQO), respectivamente.
Deficiencias en estas enzimas (ETF o ETFDH) conducen a una disminución en la FAD oxidada, la cual se torna no disponible para las reacciones de deshidrogenación dependientes de FAD, incluyendo el primer paso en la β-oxidación — un principal proceso catabólico de los ácidos grasos que tiene lugar en la mitocondria. Un defecto en la β-oxidación de ácidos grasos causa acumulación de lípidos en el musculo esquelético, llevando a una miopatía de almacenamiento de lípidos caracterizada por dolor y debilidad muscular e intolerancia al ejercicio. Junto con una dieta baja en grasas y alta en carbohidratos, la suplementación con riboflavina ha llevado a mejoramientos clínicos significantes en pacientes con mutaciones de la ETFDH. El tipo específico de mutación en ETF/ETFDH contribuye a la edad de inicio, gravedad y capacidad de respuesta al tratamiento con riboflavina (58). Adicionalmente, el reciente reporte de un hombre de 20 años de edad con una MADD sensible a la riboflavina falló en encontrar mutaciones en los genes de la ETF y ETFDH, sugiriendo que otros sitios de mutación no deben ser excluidos (57). Finalmente, deficiencias secundarias en la cadena respiratoria son observadas en MADD y parecen responder favorablemente a la suplementación con riboflavina (58, 59).

[Figura 3 - Clic para Agrandar]
Deficiencia de acil CoA-deshidrogenasa 9 (ACAD9)
Mutaciones recesivas en el gen de la ACAD9 que codifica para la acil CoA deshidrogenasa dependiente de FAD fueron encontradas en pacientes con deficiencia del complejo I mitocondrial, un trastorno de la cadena respiratoria (60). La deficiencia de ACAD9 no ha sido ligada a defectos de la β-oxidación de ácidos grasos, sugiriendo en cambio un papel en el ensamblaje del complejo I para la ACAD9 (61). El complejo I transporta electrones de la NADH a la Coenzima Q10 en la cadena transportadora de electrones. La fosforilación oxidativa defectuosa (síntesis de ATP por la cadena respiratoria) debido a la deficiencia del complejo I ha sido ligada a una amplia variedad de manifestaciones clínicas desde muerte neonatal hasta la aparición tardía de enfermedades neurodegenerativas. Los síntomas de la deficiencia de complejo I debido a mutaciones en ACAD9 incluyen debilidad muscular, intolerancia al ejercicio, acidosis láctica, encefalopatía, y cardiomiopatía. La suplementación con riboflavina (100-300 mg/día) incremento la actividad del complejo I en pacientes con formas clínicas de la deficiencia de ACAD9 que aparecieron en la infancia. Mejoras en la resistencia muscular y tolerancia al ejercicio han sido también asociadas con la suplementación de riboflavina (62-64).
Trastornos asociados con el transporte defectuoso de la riboflavina
Los genes SLC52A1, SLC52A3, y SLC52A2 codifican para los transportadores de riboflavina humanos hRTF1, hRTF2, y hRTF3, respectivamente. Mutaciones en estos genes han sido ligados con el síndrome de Brown-Vialetto-Van Laere (BVVL), un desorden neurodegenerativo raro caracterizado por su aparición de edades variable. El síndrome incluye parálisis bulbar con hipotonía y debilidad facial, sordera neurosensorial, e insuficiencia respiratoria. La falta de pérdida auditiva en la descripción clínica de los síntomas del BVVL es conocida como síndrome de Fazio-Londe (FL). Las características transitorias clínicas y bioquímicas de MADD fueron descritas en un recién nacido de una madre con deficiencia de riboflavina; esta leve deficiencia, causada por una mutación en el hRTF1, fue corregida rápidamente por la suplementación con riboflavina (65, 66). Una reciente revisión de la literatura, la cual analizó reportes de 74 pacientes afectados por el síndrome BVVL/FL, encontró que 8 de los 13 pacientes a los que se les dio riboflavina suplementaria (dosis promedio de 10 mg/kg/día) experimentaron mejoras clínicas, y los individuos tratados tuvieron una tasa de supervivencia del 100% (67). La riboflavina también restauró los niveles normales de flavina y acilcarnitina en pacientes que presentaban perfiles anormales.
Trimetilaminuria sensible a la riboflavina
La trimetilaminuria primaria es causada por la oxidación defectuosa de la trimetilamina por una flavoproteina del hígado llamada flavina monooxigenasa 3 (FMO3). Individuos con deficiencia de FMO3 tienen niveles incrementados de trimetilamina en la orina, sudor, y aliento (68). Esta condición socialmente angustiante es conocida como "síndrome de olor a pescado" debido al olor a pescado y naturaleza volátil de la trimetilamina. Mutaciones del gen FMO3 son usualmente asociadas con trimetilaminuria leve o intermitente; la condición es a veces limitada a periodos perimenstruales en sujetos femeninos o al consumo de alimentos ricos en trimetilamina. El manejo clínico de esta condición incluye restricciones dietarías de trimetilamina y sus precursores, como los alimentos ricos en colina y los mariscos, como también los vegetales crucíferos que contienen ambos precursores de la trimetilamina y antagonistas de la FMO3 (69). El uso de suplementos de riboflavina fue recientemente reportado en un paciente femenino de 17 años de edad afectado por homocistinuria no responsiva a la piridoxina (70). La enfermedad fue inicialmente tratada con betaína (un derivado de la colina), la cual causó olor corporal secundario a la deficiencia de FMO3. La suplementación con riboflavina (200 mg/día) redujo la excreción de trimetilamina y el olor corporal relacionado al tratamiento con betaína. Estos datos sugieren que la riboflavina podría ayudar a maximizar la actividad residual de la enzima FMO3 en pacientes con trimetilaminuria primaria.
Hipertensión
Aunque la etiología de la hipertensión no es clara, el polimorfismo C677T del gen de la metilentetrahidrofolato reductasa (MTHFR) es el principal determinante de las concentraciones de homocisteína y ha sido relacionado a una presión sanguínea elevada (un marcador de hipertensión) (71) y un riesgo incrementado de enfermedad coronaria cardíaca y accidente vascular (72-74). Dado que esta variante genética lleva a una actividad de la MTHFR disminuida, individuos con el genotipo 677TT podrían beneficiarse de la suplementación con riboflavina. En un ensayo inicial aleatorio, doble ciego, controlado con placebo en 77 sujetos sanos que habían sido pre-cribados para el genotipo de la MTHFR, la suplementación con riboflavina (1.6 mg/día por 12 semanas) disminuyó los niveles de homocisteína en el grupo con el genotipo 677TT MTHFR, pero en no en los grupos 677CC y 677CT de la misma edad que exhibieron niveles normales al inicio del estudio (75). Dos ensayos aleatorios, doble ciegos, controlados con placebo subsecuentes investigaron la posibilidad de la modulación de la hipertensión por la riboflavina en pacientes con enfermedades cardiovasculares (ECV) prematuras (pre-cribados para el polimorfismo MTHFR 677C→T) (76, 77). Los resultados mostraron una disminución significante de la presión sanguínea solo en pacientes con el genotipo 677TT suplementados con riboflavina (1.6 mg/día por 16 semanas) en comparación al placebo, tanto en la examinación inicial (69) como cuando la misma cohorte de pacientes en alto riesgo de ECV fue investigada nuevamente cuatro años después del ensayo original (70). Otro estudio investigó el efecto de la riboflavina en 88 pacientes hipertensos (pero sin ECV presentes) con el genotipo 677TT de la MTHFR, la mayoría de los cuales habían sido tratados con terapia antihipertensiva. Al inicio del estudio, el 60% de los participantes habían fallado en lograr el objetivo de los niveles de presión sanguínea (≤140/90 mm Hg), a pesar de haber tomado tres o más medicamentos antihipertensivos. La reducción en la presión sanguínea después de la suplementación con riboflavina (1.6 mg/día por 16 semanas) en estos pacientes sugirió que el exceso del riesgo de hipertensión ligado a esta variación genética pudo ser superado al optimizar el estatus de la riboflavina (78).
Cáncer
Los agentes anticancerosos frecuentemente muestran varios efectos secundarios que pueden forzar a los pacientes a limitar la dosis o a descontinuar el tratamiento. El efecto antioxidante de la coadministración de riboflavina (10 mg/día), niacina (50 mg/día), y coenzima Q10 (100 mg/día) fue evaluado en 78 pacientes postmenopáusicos con cáncer de seno tratados con tamoxifeno por 90 días. Esta suplementación previno efectivamente el estrés oxidativo asociado con el tratamiento con tamoxifeno (79). La riboflavina puede también actuar como un fotosensibilizador, y esta propiedad pudiese ser de valor en la terapia fotodinámica del cáncer. Un modelo de ratón fue usado para evaluar el efecto de la riboflavina en combinación con cisplatino, uno de los agentes anticancerosos más efectivos. Bajo exposición a la luz, la administración de riboflavina redujo el daño al ADN inducido por el cisplatino en el hígado y los riñones (80). Estos resultados son prometedores, pero estudios en humanos son necesarios para examinar si la riboflavina podría ser un adjunto para la quimioterapia.
Trastornos de la córnea
La ectasia corneal es una condición del ojo caracterizada por irregularidades de la córnea que afectan la visión. La reticulación corneal — un nuevo procedimiento usado por profesionales para limitar la progresión del daño corneal — involucra el uso de riboflavina en conjunción con irradiación de luz ultravioleta. La reticulación corneal modifica las propiedades de la córnea y fortalece su arquitectura (81).
Fuentes
Fuentes alimenticias
La mayoría de los alimentos derivados de plantas y animales contienen al menos pequeñas cantidades de riboflavina. En los EE.UU. se ha enriquecido la harina de trigo y el pan con riboflavina (y también con tiamina, niacina, y hierro) desde 1943. Datos provenientes de encuestas dietarías masivas indican que la ingesta promedio de riboflavina en hombres es de alrededor de 2 mg/día, y en mujeres es alrededor de 1.5 mg/día; ambas ingestas están muy por encima de la IDR. Los niveles de ingesta fueron similares para una población de hombres y mujeres de la tercera edad (1). La riboflavina es fácilmente destruida luego de su exposición a la luz. Por ello, hasta un 50% de la riboflavina de la leche contenida en una botella de vidrio transparente puede destruirse luego de dos horas de exposición a luz solar intensa (7). Algunos alimentos con cantidades substanciales de riboflavina están listados en la Tabla 2, junto con su contenido de riboflavina en miligramos (mg). Para mayor información sobre el contenido de nutrientes de los alimentos, busque la base de datos de composición de los alimentos del USDA.
| Alimento | Porción | Riboflavina (mg) |
|---|---|---|
| Cereal fortificado | 1 taza | 0.59 to 2.27 |
| Leche (descremada) | 1 taza (8 onzas) | 0.34 |
| Queso cheddar | 1 onza | 0.11 |
| Huevo (cocido) | 1 grande | 0.27 |
| Almendras | 1 onza | 0.23 |
| Salmón (cocido) | 3 onzas* | 0.12 |
| Rodaballo (a la parrilla) | 3 onzas | 0.08 |
| Pollo, carne blanca (asada) | 3 onzas | 0.08 |
| Pollo, carne oscura (asada) | 3 onzas | 0.16 |
| Carne de vacuno (cocido) | 3 onzas | 0.16 |
| Brócoli (hervido) | ½ taza, picado | 0.10 |
| Espárragos (hervido) | 6 varitas | 0.13 |
| Espinaca (hervido) | ½ taza | 0.21 |
| Pan, trigo entero | 1 rebanada | 0.06 |
| Pan, blanco (enriquecido) | 1 rebanada | 0.08 |
| *Una porción de tres onzas de carnes es del tamaño de una baraja de cartas. | ||
Suplementos
Las formas más comunes de riboflavina disponible en los suplementos son riboflavina y riboflavina-5’-monofosfato. La riboflavina se encuentra con frecuencia en preparaciones multivitamínicas y de vitaminas del complejo B (26).
Seguridad
Toxicidad
Se desconocen efectos tóxicos o adversos debido a ingestas elevadas de riboflavina en humanos. Estudios en células de cultivo señalan que un exceso de riboflavina podría aumentar el riesgo de ruptura de hebras de ADN en presencia de cromio (IV), un conocido carcinógeno (27). Esto podría ser motivo de preocupación para trabajadores expuestos al cromo, pero no hay información disponible de estudios en humanos. Se ha encontrado que la terapia con dosis elevadas de riboflavina intensifica el color de la orina a un amarillo fuerte (flavinuria), pero este es un efecto secundario inocuo. La Junta de Nutrición y Alimentos no estableció un nivel máximo de ingesta tolerable (NM) cuando se revisó la IDR en 1998 (1).
Interacción con drogas
Varios estudios preliminares señalaron que las mujeres tomando anticonceptivos orales (AO) en dosis altas habían disminuido su estado nutricional de riboflavina. Sin embargo, cuando los investigadores controlaron la ingesta de riboflavina alimentaria, no encontraron diferencias entre las usuarias de AO y las no usuarias (1). Los derivados de la fenotiazina (como el medicamento antipsicótico clorpromazina y los antidepresivos tricíclicos) inhiben la incorporación de riboflavina en el FAD y el FMN, así como lo hace la quinacrina, un medicamento contra la malaria, y la adriamicina, un agente quimioterapéutico contra el cáncer (4). La utilización a largo plazo del anticonvulsivo fenobarbital podría incrementar la destrucción de riboflavina por parte de las enzimas hepáticas, aumentando el riesgo de una deficiencia (3).
Recomendación del LPI
La IDR de riboflavina (1.3 mg/día para hombres y 1.1 mg/día para mujeres), que debiera prevenir una deficiencia en la mayoría de las personas, se alcanza fácilmente consumiendo una dieta variada. Consumir una dieta variada debería proporcionar 1.5 mg a 2 mg de riboflavina al día. Siguiendo la recomendación del Instituto Linus Pauling (LPI) de tomar un suplemento multivitamínico/multimineral que contenga el 100% del Valor Diario (VD) asegurará una ingesta de al menos 1.7 mg de riboflavina al día.
Adultos mayores (>50 años)
Algunos expertos en nutrición y envejecimiento consideraron que la IDR (1.3 mg/día para hombres y 1.1 mg/día para mujeres) dejaba un pequeño margen de error en personas sobre los 50 años de edad (28, 29). Un estudio reciente en personas que viven de manera independiente entre 65 y 90 años de edad reveló que casi el 25% consumía menos riboflavina que la ingesta recomendada, y que un 10% presentaba evidencia bioquímica de una deficiencia (30). Adicionalmente, estudios epidemiológicos de prevalencia de cataratas señalaron que la ingesta de riboflavina de 1.6 a 2.2 mg/día podría reducir el riesgo de sufrir de cataratas relacionadas con la edad. En individuos donde sus dietas podrían no proporcionar riboflavina suficiente, especialmente en aquellos sobre 50 años, deberían considerar consumir un suplemento multivitamínico/multimineral, el cual aporta generalmente, al menos 1.7 mg de riboflavina al día.
Autores y Críticos
Escrito en 2000 por:
Jane Higdon, Ph.D.
Instituto Linus Pauling
Universidad Estatal de Oregon
Actualizado en Septiembre de 2002 por:
Jane Higdon, Ph.D.
Instituto Linus Pauling
Universidad Estatal de Oregon
Actualizado en Junio de 2007 por:
Victoria J. Drake, Ph.D.
Instituto Linus Pauling
Universidad Estatal de Oregon
Actualizado en Julio de 2013 por:
Barbara Delage, Ph.D.
Instituto Linus Pauling
Universidad Estatal de Oregon
Revisado en Diciembre de 2013 por:
Helene McNulty, Ph.D., R.D.
Profesor de Nutrición Humana y Dietética
Centro para la Alimentación y la Salud de Irlanda del Norte (NICHE)
Universidad de Ulster
Coleraine, Reino Unido
Revisado en Diciembre de 2013 por:
Adrian McCann, Ph.D.
Asociado en Investigación
Centro para la Alimentación y la Salud de Irlanda del Norte (NICHE)
Universidad de Ulster
Coleraine, Reino Unido
Traducido al Español en 2016 por:
Silvia Vazquez Lima
Instituto Linus Pauling
Universidad Estatal de Oregon
Originalmente traducido al español en 2012 por Guillermo Sandoval y editado por Andrew Quest (Ph.D.) y Lisette Leyton (Ph.D.), todos provenientes de la Universidad de Chile. Estos esfuerzos fueron patrocinados por el projecto Anillo #ACT1111, CONICYT-Chile, programa PIA.
Derechos de autoría 2000-2026 Instituto Linus Pauling
Figure Alternative Text
Figure 1. Glutathione oxidation-reduction (redox) cycle
The figure shows the glutathione oxidation-reduction (redox) cycle. Riboflavin, in the form of flavin adenine dinucleotide (FAD), is a required cofactor for glutathione reductase. This enzyme reduces GSSG (oxidized glutathione) to two molecules of glutathione (GSH) using NADPH and a proton (H+), releasing NADP+. The figure also shows that the selenium-dependent enzyme glutathione peroxidase oxidizes two molecules of glutathione back to GSSG while converting one molecule of hydrogen peroxide (H2O2) into two molecules of water (H2O).
Figure 2. Folate and nucleic acid metabolism
The figure highlights that riboflavin in the form of flavin adenine dinucleotide (FAD) is a required cofactor for the enzyme methylenetetrahydrofolate reductase (MTHFR), which reduces 5,10-methylenetetrahydrofolate to 5-methyltetrahydrofolate. Methionine synthase, a vitamin B12-dependent enzyme, converts homocysteine to methionine, regenerating tetrahydrofolate (THF) from 5-methylTHF. Methionine in the form of S-adenosylmethionine (SAM) is required for many methylation reactions, including DNA methylation.
Figure 3. Fatty acid β-oxidation and the electron transfer flavoprotein system
The figure shows a pathway of fatty acid β-oxidation and the electron transfer flavoprotein (ETF) system. Highlighted are the flavoproteins essential for electron transfer from the fatty acid β-oxidation pathway to the respiratory chain.
At the top, acyl-carnitine and coenzyme A are converted to carnitine and acyl-CoA. Acyl-CoA then enters β-oxidation: acyl-CoA dehydrogenase catalyzes the conversion of acyl-CoA to trans-delta2-enoyl-CoA. This enzyme requires riboflavin in the form of flavin adenine dinucleotide (FAD) and reduces FAD to FADH2 during the reaction.
Electrons from FADH2 are transferred to ETF, which passes electrons to the ETF-ubiquinone oxidoreductase system (ETF-QO) in the respiratory chain through iron-sulfur clusters, ultimately reducing ubiquinone to ubiquinol.
The figure shows further steps of β-oxidation: Trans-delta2-enoyl-CoA is converted to β-ketoacyl-CoA, then to acetyl-CoA and a shortened acyl-CoA (by two carbons) following condensation with coenzyme A.
The figure legend notes that multiple acyl-CoA dehydrogenase deficiency (MADD) results from mutations in the genes coding for the ETF or ETF-QO.
Referencias
- Food and Nutrition Board, Institute of Medicine. Riboflavin. Dietary Reference Intakes: Thiamin, Riboflavin, Niacin, Vitamin B6, Vitamin B12, Pantothenic Acid, Biotin, and Choline. Washington D.C.: National Academy Press; 1998:87-122. (National Academy Press)
- Brody T. Nutritional Biochemistry. 2nd ed. San Diego: Academic Press; 1999.
- McCormick DB. Riboflavin. In: Shils M, Olson JA, Shike M, Ross AC, eds. Modern Nutrition in Health and Disease. 9th ed. Baltimore: Williams & Wilkins; 1999:391-399.
- Powers HJ. Current knowledge concerning optimum nutritional status of riboflavin, niacin and pyridoxine. Proc Nutr Soc. 1999;58(2):435-440. (PubMed)
- Rivlin RS. Riboflavin. In: Ziegler EE, Filer LJ, eds. Present Knowledge in Nutrition. 7th ed. Washington D.C.: ILSI Press; 1996:167-173.
- Hoey L, McNulty H, Strain JJ. Studies of biomarker responses to intervention with riboflavin: a systematic review. Am J Clin Nutr. 2009;89(6):1960S-1980S. (PubMed)
- Bohles H. Antioxidative vitamins in prematurely and maturely born infants. Int J Vitam Nutr Res. 1997;67(5):321-328. (PubMed)
- McCormick DB. Two interconnected B vitamins: riboflavin and pyridoxine. Physiol Rev. 1989;69(4):1170-1198. (PubMed)
- Madigan SM, Tracey F, McNulty H, et al. Riboflavin and vitamin B-6 intakes and status and biochemical response to riboflavin supplementation in free-living elderly people. Am J Clin Nutr. 1998;68(2):389-395. (PubMed)
- Lowik MR, van den Berg H, Kistemaker C, Brants HA, Brussaard JH. Interrelationships between riboflavin and vitamin B6 among elderly people (Dutch Nutrition Surveillance System). Int J Vitam Nutr Res. 1994;64(3):198-203. (PubMed)
- Jacques PF, Bostom AG, Wilson PW, Rich S, Rosenberg IH, Selhub J. Determinants of plasma total homocysteine concentration in the Framingham Offspring cohort. Am J Clin Nutr. 2001;73(3):613-621. (PubMed)
- Jacques PF, Kalmbach R, Bagley PJ, et al. The relationship between riboflavin and plasma total homocysteine in the Framingham Offspring cohort is influenced by folate status and the C677T transition in the methylenetetrahydrofolate reductase gene. J Nutr. 2002;132(2):283-288. (PubMed)
- Powers HJ, Weaver LT, Austin S, Beresford JK. A proposed intestinal mechanism for the effect of riboflavin deficiency on iron loss in the rat. Br J Nutr. 1993;69(2):553-561. (PubMed)
- Powers HJ, Hill MH, Mushtaq S, Dainty JR, Majsak-Newman G, Williams EA. Correcting a marginal riboflavin deficiency improves hematologic status in young women in the United Kingdom (RIBOFEM). Am J Clin Nutr. 2011;93(6):1274-1284. (PubMed)
- Powers HJ. Riboflavin-iron interactions with particular emphasis on the gastrointestinal tract. Proc Nutr Soc. 1995;54(2):509-517. (PubMed)
- Kalaivani K. Prevalence & consequences of anaemia in pregnancy. Indian J Med Res. 2009;130(5):627-633. (PubMed)
- Worldwide prevalence of anaemia 1993-2005: WHO global database on anaemia. de Benoist B, McLean E, Egli I, Cogswell M, eds. 2008; World Health Organization Press. Available at: http://www.who.int/nutrition/publications/micronutrients/anaemia_iron_deficiency/9789241596657/en/. Accessed 7/22/13.
- Pena-Rosas JP, Viteri FE. Effects of routine oral iron supplementation with or without folic acid for women during pregnancy. Cochrane Database Syst Rev. 2006(3):CD004736. (PubMed)
- Suprapto B, Widardo, Suhanantyo. Effect of low-dosage vitamin A and riboflavin on iron-folate supplementation in anaemic pregnant women. Asia Pac J Clin Nutr. 2002;11(4):263-267. (PubMed)
- Ma AG, Schouten EG, Zhang FZ, et al. Retinol and riboflavin supplementation decreases the prevalence of anemia in Chinese pregnant women taking iron and folic Acid supplements. J Nutr. 2008;138(10):1946-1950. (PubMed)
- Crombleholme WR. Obstetrics. In: Tierney LM, McPhee SJ, Papadakis MA, eds. Current Medical Treatment and Diagnosis. 37th ed. Stamford: Appleton and Lange; 1998:731-734.
- Wacker J, Fruhauf J, Schulz M, Chiwora FM, Volz J, Becker K. Riboflavin deficiency and preeclampsia. Obstet Gynecol. 2000;96(1):38-44. (PubMed)
- Wang XM, Wu HY, Qiu XJ. Methylenetetrahydrofolate reductase (MTHFR) gene C677T polymorphism and risk of preeclampsia: an updated meta-analysis based on 51 studies. Arch Med Res. 2013;44(3):159-168. (PubMed)
- Xia XP, Chang WW, Cao YX. Meta-analysis of the methylenetetrahydrofolate reductase C677T polymorphism and susceptibility to pre-eclampsia. Hypertens Res. 2012;35(12):1129-1134. (PubMed)
- Braekke K, Ueland PM, Harsem NK, Karlsen A, Blomhoff R, Staff AC. Homocysteine, cysteine, and related metabolites in maternal and fetal plasma in preeclampsia. Pediatr Res. 2007;62(3):319-324. (PubMed)
- Neugebauer J, Zanre Y, Wacker J. Riboflavin supplementation and preeclampsia. Int J Gynaecol Obstet. 2006;93(2):136-137. (PubMed)
- Heese P, Linnebank M, Semmler A, et al. Alterations of homocysteine serum levels during alcohol withdrawal are influenced by folate and riboflavin: results from the German Investigation on Neurobiology in Alcoholism (GINA). Alcohol Alcohol. 2012;47(5):497-500. (PubMed)
- Soares MJ, Satyanarayana K, Bamji MS, Jacob CM, Ramana YV, Rao SS. The effect of exercise on the riboflavin status of adult men. Br J Nutr. 1993;69(2):541-551. (PubMed)
- Ruston D, Hoare J, Henderson L, et al. The National Diet & Nutrition Survey: adults aged 19 to 64 years: Nutritional status (anthropometry and blood analytes), blood pressure and physical activity. Volume 4. London: The Stationary Office; 2004.
- Mares-Perlman JA, Brady WE, Klein BE, et al. Diet and nuclear lens opacities. Am J Epidemiol. 1995;141(4):322-334. (PubMed)
- Leske MC, Wu SY, Hyman L, et al. Biochemical factors in the lens opacities. Case-control study. The Lens Opacities Case-Control Study Group. Arch Ophthalmol. 1995;113(9):1113-1119. (PubMed)
- Cumming RG, Mitchell P, Smith W. Diet and cataract: the Blue Mountains Eye Study. Ophthalmology. 2000;107(3):450-456. (PubMed)
- Hankinson SE, Stampfer MJ, Seddon JM, et al. Nutrient intake and cataract extraction in women: a prospective study. BMJ. 1992;305(6849):335-339. (PubMed)
- Jacques PF, Taylor A, Moeller S, et al. Long-term nutrient intake and 5-year change in nuclear lens opacities. Arch Ophthalmol. 2005;123(4):517-526. (PubMed)
- McNulty H, Strain JJ, Pentieva K, Ward M. C(1) metabolism and CVD outcomes in older adults. Proc Nutr Soc. 2012;71(2):213-221. (PubMed)
- Holmes MV, Newcombe P, Hubacek JA, et al. Effect modification by population dietary folate on the association between MTHFR genotype, homocysteine, and stroke risk: a meta-analysis of genetic studies and randomised trials. Lancet. 2011;378(9791):584-594. (PubMed)
- Wilcken B, Bamforth F, Li Z, et al. Geographical and ethnic variation of the 677C>T allele of 5,10 methylenetetrahydrofolate reductase (MTHFR): findings from over 7000 newborns from 16 areas world wide. J Med Genet. 2003;40(8):619-625. (PubMed)
- McGlynn AP, Wasson GR, O'Reilly SL, et al. Low colonocyte folate is associated with uracil misincorporation and global DNA hypomethylation in human colorectum. J Nutr. 2013;143(1):27-33. (PubMed)
- Guenther BD, Sheppard CA, Tran P, Rozen R, Matthews RG, Ludwig ML. The structure and properties of methylenetetrahydrofolate reductase from Escherichia coli suggest how folate ameliorates human hyperhomocysteinemia. Nat Struct Biol. 1999;6(4):359-365. (PubMed)
- Yin G, Ming H, Zheng X, Xuan Y, Liang J, Jin X. Methylenetetrahydrofolate reductase C677T gene polymorphism and colorectal cancer risk: A case-control study. Oncol Lett. 2012;4(2):365-369. (PubMed)
- Gao S, Ding LH, Wang JW, Li CB, Wang ZY. Diet folate, DNA methylation and polymorphisms in methylenetetrahydrofolate reductase in association with the susceptibility to gastric cancer. Asian Pac J Cancer Prev. 2013;14(1):299-302. (PubMed)
- Wen YY, Yang SJ, Zhang JX, Chen XY. Methylenetetrahydrofolate reductase genetic polymorphisms and esophageal squamous cell carcinoma susceptibility: a meta-analysis of case-control studies. Asian Pac J Cancer Prev. 2013;14(1):21-25. (PubMed)
- Powers HJ, Hill MH, Welfare M, et al. Responses of biomarkers of folate and riboflavin status to folate and riboflavin supplementation in healthy and colorectal polyp patients (the FAB2 Study). Cancer Epidemiol Biomarkers Prev. 2007;16(10):2128-2135. (PubMed)
- Zschabitz S, Cheng TY, Neuhouser ML, et al. B vitamin intakes and incidence of colorectal cancer: results from the Women's Health Initiative Observational Study cohort. Am J Clin Nutr. 2013;97(2):332-343. (PubMed)
- Kennedy DA, Stern SJ, Matok I, et al. Folate Intake, MTHFR Polymorphisms, and the Risk of Colorectal Cancer: A Systematic Review and Meta-Analysis. J Cancer Epidemiol. 2012:952508. doi: 10.1155/2012/952508. (PubMed)
- He Y, Ye L, Shan B, Song G, Meng F, Wang S. Effect of riboflavin-fortified salt nutrition intervention on esophageal squamous cell carcinoma in a high incidence area, China. Asian Pac J Cancer Prev. 2009;10(4):619-622. (PubMed)
- Bassett JK, Hodge AM, English DR, et al. Dietary intake of B vitamins and methionine and risk of lung cancer. Eur J Clin Nutr. 2012;66(2):182-187. (PubMed)
- Bassett JK, Baglietto L, Hodge AM, et al. Dietary intake of B vitamins and methionine and breast cancer risk. Cancer Causes Control. 2013;24(8):1555-1563. (PubMed)
- Bassett JK, Severi G, Hodge AM, et al. Dietary intake of B vitamins and methionine and prostate cancer incidence and mortality. Cancer Causes Control. 2012;23(6):855-863. (PubMed)
- Qiao YL, Dawsey SM, Kamangar F, et al. Total and cancer mortality after supplementation with vitamins and minerals: follow-up of the Linxian General Population Nutrition Intervention Trial. J Natl Cancer Inst. 2009;101(7):507-518. (PubMed)
- Schoenen J, Jacquy J, Lenaerts M. Effectiveness of high-dose riboflavin in migraine prophylaxis. A randomized controlled trial. Neurology. 1998;50(2):466-470. (PubMed)
- Sandor PS, Afra J, Ambrosini A, Schoenen J. Prophylactic treatment of migraine with β-blockers and riboflavin: differential effects on the intensity dependence of auditory evoked cortical potentials. Headache. 2000;40(1):30-35. (PubMed)
- Boehnke C, Reuter U, Flach U, Schuh-Hofer S, Einhaupl KM, Arnold G. High-dose riboflavin treatment is efficacious in migraine prophylaxis: an open study in a tertiary care centre. Eur J Neurol. 2004;11(7):475-477. (PubMed)
- Maizels M, Blumenfeld A, Burchette R. A combination of riboflavin, magnesium, and feverfew for migraine prophylaxis: a randomized trial. Headache. 2004;44(9):885-890. (PubMed)
- 55. MacLennan SC, Wade FM, Forrest KM, Ratanayake PD, Fagan E, Antony J. High-dose riboflavin for migraine prophylaxis in children: a double-blind, randomized, placebo-controlled trial. J Child Neurol. 2008;23(11):1300-1304. (PubMed)
- Bruijn J, Duivenvoorden H, Passchier J, Locher H, Dijkstra N, Arts WF. Medium-dose riboflavin as a prophylactic agent in children with migraine: a preliminary placebo-controlled, randomised, double-blind, cross-over trial. Cephalalgia. 2010;30(12):1426-1434. (PubMed)
- Cotelli MS, Vielmi V, Rimoldi M, et al. Riboflavin-responsive multiple acyl-CoA dehydrogenase deficiency with unknown genetic defect. Neurol Sci. 2012;33(6):1383-1387. (PubMed)
- Olsen RK, Olpin SE, Andresen BS, et al. ETFDH mutations as a major cause of riboflavin-responsive multiple acyl-CoA dehydrogenation deficiency. Brain. 2007;130(Pt 8):2045-2054. (PubMed)
- Liang WC, Ohkuma A, Hayashi YK, et al. ETFDH mutations, CoQ10 levels, and respiratory chain activities in patients with riboflavin-responsive multiple acyl-CoA dehydrogenase deficiency. Neuromuscul Disord. 2009;19(3):212-216. (PubMed)
- Haack TB, Danhauser K, Haberberger B, et al. Exome sequencing identifies ACAD9 mutations as a cause of complex I deficiency. Nat Genet. 2010;42(12):1131-1134. (PubMed)
- Nouws J, Nijtmans L, Houten SM, et al. Acyl-CoA dehydrogenase 9 is required for the biogenesis of oxidative phosphorylation complex I. Cell Metab. 2010;12(3):283-294. (PubMed)
- Scholte HR, Busch HF, Bakker HD, Bogaard JM, Luyt-Houwen IE, Kuyt LP. Riboflavin-responsive complex I deficiency. Biochim Biophys Acta. 1995;1271(1):75-83. (PubMed)
- Gerards M, van den Bosch BJ, Danhauser K, et al. Riboflavin-responsive oxidative phosphorylation complex I deficiency caused by defective ACAD9: new function for an old gene. Brain. 2011;134(Pt 1):210-219. (PubMed)
- Garone C, Donati MA, Sacchini M, et al. Mitochondrial Encephalomyopathy Due to a Novel Mutation in ACAD9. JAMA Neurol. 2013:1-3. (PubMed)
- Chiong MA, Sim KG, Carpenter K, et al. Transient multiple acyl-CoA dehydrogenation deficiency in a newborn female caused by maternal riboflavin deficiency. Mol Genet Metab. 2007;92(1-2):109-114. (PubMed)
- Ho G, Yonezawa A, Masuda S, et al. Maternal riboflavin deficiency, resulting in transient neonatal-onset glutaric aciduria Type 2, is caused by a microdeletion in the riboflavin transporter gene GPR172B. Hum Mutat. 2011;32(1):E1976-1984. (PubMed)
- Bosch AM, Stroek K, Abeling NG, Waterham HR, Ijlst L, Wanders RJ. The Brown-Vialetto-Van Laere and Fazio Londe syndrome revisited: natural history, genetics, treatment and future perspectives. Orphanet J Rare Dis. 2012;7:83. doi: 10.1186/1750-1172-7-83. (PubMed)
- Mackay RJ, McEntyre CJ, Henderson C, Lever M, George PM. Trimethylaminuria: causes and diagnosis of a socially distressing condition. Clin Biochem Rev. 2011;32(1):33-43. (PubMed)
- Phillips IR, Shephard EA. Trimethylaminuria. 2007 Oct 8 [Updated 2011 Apr 19]. In: Pagon RA, Adam MP, Bird TD, et al., editors. GeneReviews™ [Internet]. Seattle: University of Washington, Seattle; 1993-2013. Available at: http://www.ncbi.nlm.nih.gov/books/NBK1103/.
- Manning NJ, Allen EK, Kirk RJ, Sharrard MJ, Smith EJ. Riboflavin-responsive trimethylaminuria in a patient with homocystinuria on betaine therapy. JIMD Rep. 2012;5:71-75. (PubMed)
- Niu WQ, You YG, Qi Y. Strong association of methylenetetrahydrofolate reductase gene C677T polymorphism with hypertension and hypertension-in-pregnancy in Chinese: a meta-analysis. J Hum Hypertens. 2012;26(4):259-267. (PubMed)
- Almawi WY, Khan A, Al-Othman SS, Bakhiet M. Case-control Study of methylenetetrahydrofolate reductase mutations and hyperhomocysteinemia and risk of stroke. J Stroke Cerebrovasc Dis. 2009;18(5):407-408. (PubMed)
- Heux S, Morin F, Lea RA, Ovcaric M, Tajouri L, Griffiths LR. The methylentetrahydrofolate reductase gene variant (C677T) as a risk factor for essential hypertension in Caucasians. Hypertens Res. 2004;27(9):663-667. (PubMed)
- Klerk M, Verhoef P, Clarke R, et al. MTHFR 677C→T polymorphism and risk of coronary heart disease: a meta-analysis. JAMA. 2002;288(16):2023-2031. (PubMed)
- McNulty H, Dowey le RC, Strain JJ, et al. Riboflavin lowers homocysteine in individuals homozygous for the MTHFR 677C→T polymorphism. Circulation. 2006;113(1):74-80. (PubMed)
- Horigan G, McNulty H, Ward M, Strain JJ, Purvis J, Scott JM. Riboflavin lowers blood pressure in cardiovascular disease patients homozygous for the 677C→T polymorphism in MTHFR. J Hypertens. 2010;28(3):478-486. (PubMed)
- Wilson CP, Ward M, McNulty H, et al. Riboflavin offers a targeted strategy for managing hypertension in patients with the MTHFR 677TT genotype: a 4-y follow-up. Am J Clin Nutr. 2012;95(3):766-772. (PubMed)
- Wilson CP, McNulty H, Ward M, et al. Blood pressure in treated hypertensive individuals with the MTHFR 677TT genotype is responsive to intervention with riboflavin: findings of a targeted randomized trial. Hypertension. 2013;61(6):1302-1308. (PubMed)
- Yuvaraj S, Premkumar VG, Vijayasarathy K, Gangadaran SG, Sachdanandam P. Augmented antioxidant status in Tamoxifen treated postmenopausal women with breast cancer on co-administration with Coenzyme Q10, Niacin and Riboflavin. Cancer Chemother Pharmacol. 2008;61(6):933-941. (PubMed)
- Hassan I, Chibber S, Khan AA, Naseem I. Riboflavin ameliorates cisplatin induced toxicities under photoillumination. PLoS One. 2012;7(5):e36273. (PubMed)
- Raiskup F, Spoerl E. Corneal crosslinking with riboflavin and ultraviolet A. I. Principles. Ocul Surf. 2013;11(2):65-74. (PubMed)
- Hendler SS, Rorvik DR, eds. PDR for Nutritional Supplements. Montvale: Medical Economics Company, Inc; 2001.
- Sugiyama M. Role of physiological antioxidants in chromium(VI)-induced cellular injury. Free Radic Biol Med. 1992;12(5):397-407. (PubMed)
- Subramanian VS, Subramanya SB, Ghosal A, Said HM. Chronic alcohol feeding inhibits physiological and molecular parameters of intestinal and renal riboflavin transport. Am J Physiol Cell Physiol. 2013; 305(5):C539-C546. (PubMed)
- Russell RM, Suter PM. Vitamin requirements of elderly people: an update. Am J Clin Nutr. 1993;58(1):4-14. (PubMed)
- Blumberg J. Nutritional needs of seniors. J Am Coll Nutr. 1997;16(6):517-523. (PubMed)
- Lopez-Sobaler AM, Ortega RM, Quintas ME, et al. The influence of vitamin B2 intake on the activation coefficient of erythrocyte glutation reductase in the elderly. J Nutr Health Aging. 2002;6(1):60-62. (PubMed)
- Gariballa S, Ullegaddi R. Riboflavin status in acute ischaemic stroke. Eur J Clin Nutr. 2007;61(10):1237-1240. (PubMed)
- Yazdanpanah N, Uitterlinden AG, Zillikens MC, et al. Low dietary riboflavin but not folate predicts increased fracture risk in postmenopausal women homozygous for the MTHFR 677 T allele. J Bone Miner Res. 2008;23(1):86-94. (PubMed)
Exclusión de Responsabilidades
El Centro de Informaciones sobre Micronutrientes del Instituto Linus Pauling entrega información científica relacionado con aspectos de importancia en salud pública de factores dietéticos, suplementos, comestibles y bebidas para el público en general. Esta información se entrega con el entendimiento que ni el autor ni la imprenta están entregando consejos de naturaleza médica, psicológica o nutricional. La información no debe ser usada para reemplazar la consulta con profesionales competentes en el área de cuidado de salud o de nutrición.
La información entregada con relación a factores y suplementos dietéticos, comestibles, y bebestibles, contenida en este sitio web, no cubre todos los usos, acciones, precauciones, efectos secundarios, e interacciones posibles. No debe ser considerado como consejo nutricional o médico para resolver problemas individuales. La responsabilidad legal por las acciones individuales u omisiones que se derivan del uso de este sitio está expresamente excluida.
No se puede copiar, modificar, distribuir, transmitir, realizar, publicar o vender ninguno de los materiales sujetos a derechos de autoría de este sitio web. Puede enlazar a este sitio web, pero debe incluir la siguiente declaración:
"Este enlace lleva a un sitio web proporcionado por el Instituto Linus Pauling en la Universidad Estatal de Oregon. [Su nombre] no está afiliado ni respaldado por el Instituto Linus Pauling o la Universidad Estatal de Oregon."