Resumen
- Folato es un término genérico que hace referencia tanto a los folatos naturales en los alimentos como al ácido fólico, la forma sintética usada en suplementos y alimentos fortificados. El folato es crítico en el metabolismo de los precursores del ácido nucleico y varios aminoácidos, como también en las reacciones de metilación. (Más información)
- Una severa deficiencia de folato o vitamina B12 puede conducir a una anemia megaloblástica, la cual causa fatiga, debilitamiento, y dificultad para respirar. El tratamiento inadecuado de la anemia megaloblástica dependiente de la vitamina B12 con altas dosis de ácido fólico suplementario puede potencialmente retrasar el diagnóstico de la deficiencia de vitamina B12 y así dejar al individuo en un riesgo de desarrollar un daño cerebral irreversible. (Más información)
- El estatus del folato es influenciado por la presencia de variaciones genéticas en el metabolismo del folato, particularmente aquellos encontrados en el gen 5,10-metilentetrahidrofolato reductasa (MTHFR). (Más información)
- Un estatus inadecuado de folato durante el embarazo incrementa el riesgo de anomalías congénitas. La introducción de la fortificación obligatoria con ácido fólico de productos de granos refinados en los EE. UU. en 1998 ha reducido la prevalencia de defectos del tubo neural en recién nacidos. A pesar de todo, el estatus del folato es considerado inadecuado en la mayoría de mujeres a nivel mundial en edad de procrear. Por otra parte, los factores genéticos podrían modificar el riesgo de defectos del tubo neural al incrementar la susceptibilidad a una deficiencia de folato durante el embarazo. Varios estudios están actualmente investigando el papel de la suplementación con ácido fólico en la prevención de anomalías congénitas aparte de los defectos del tubo neural. (Más información)
- La deficiencia de folato y las concentraciones de homocisteína en la sangre están asociadas con un riesgo incrementado de contraer enfermedades cardiovasculares (ECV). Aunque se ha probado que la suplementación con ácido fólico ha sido efectiva en el control de las concentraciones de homocisteína circulante, el efecto de la disminución de la homocisteína en la incidencia de las ECV es todavía debatido. (Más información)
- Un estatus bajo del folato ha sido ligado con un riesgo incrementado de cáncer. Sin embargo, ensayos de intervención con altas dosis de ácido fólico no han mostrado generalmente algún beneficio en la incidencia de cáncer. (Más información)
- Estudios de cohorte prospectivos han reportado una asociación inversa entre en el estatus del folato y el riesgo de cáncer colorrectal especialmente entre hombres. La relación entre el estatus del folato y el riesgo de cáncer es sin embargo complejo y requiere de más investigación. (Más información)
- El folato es esencial para el desarrollo y función del cerebro. Un estatus bajo de folato y/o altas concentraciones de homocisteína se asocian con una disfunción cognitiva en el envejecimiento (desde deterioros leves hasta demencia). Aún se desconoce si las vitaminas B suplementarias, incluyendo el ácido fólico, tendrán beneficios a largo plazo en el mantenimiento de la salud cognitiva. (Más información)
- Varios trastornos autosómicos recesivos que afectan el transporte y metabolismo del folato pueden ser tratados con altas dosis de ácido folínico un derivado del folato. (Más información)

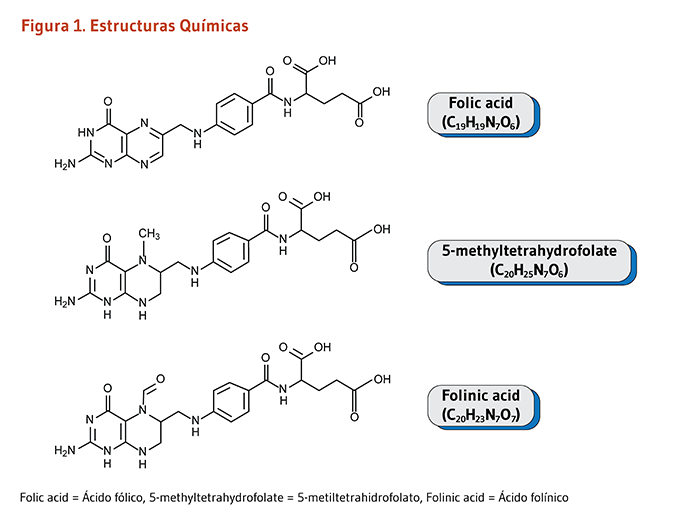
Los términos de folato y el ácido fólico se usan indistintamente para esta vitamina B soluble en agua, que también es conocida como vitamina B9 o folacina. Los folatos producidos naturalmente existen en muchas formas químicas; los folatos se encuentran en los alimentos, como también en formas metabólicamente activas en cuerpo humano. El ácido fólico es la mayor forma sintética encontrada en alimentos fortificados y suplementos vitamínicos. Otras formas sintéticas incluyen ácido folínico (Figura 1) y ácido levomefólico. El ácido fólico no tiene alguna actividad biológica a menos que sea convertido en folatos (1). En la siguiente discusión, las formas encontradas en alimentos o en el cuerpo son referidos como “folatos,” mientras que la forma encontrada en suplementos o alimentos fortificados es referida como “ácido fólico.”
Función
Metabolismo de un carbono
La única función de las coenzimas del folato en el cuerpo es aparentemente mediar la transferencia de unidades de un carbono (2). Las coenzimas del folato actúan como aceptores y dadores de unidades de un carbono en una variedad de reacciones críticas para el metabolismo de los ácidos nucleicos y los aminoácidos (Figura 2) (3).

Metabolismo de los ácidos nucleicos
Las coenzimas folato juegan un papel vital en el metabolismo del ADN a través de dos diferentes vías. (1) La síntesis de ADN desde sus precursores (timidinas y purinas) depende de las coenzimas folato. (2) Para la síntesis de metionina de la homocisteína se requiere de una coenzima folato, y la metionina es necesaria para la síntesis de S-adenosilmetionina (SAM). SAM es un dador de grupos metilo (una unidad de carbono) usado en muchas reacciones biológicas de metilación, incluyendo la metilación de una serie de sitios en el ADN, ARN, proteínas, y fosfolípidos. La metilación del ADN juega un importante papel en el control de la expresión de genes y es crítica durante la diferenciación celular. Aberraciones en la metilación del ADN han sido ligadas al desarrollo de cáncer (véase Cáncer).
Metabolismo de los aminoácidos
Las coenzimas del folato son requeridas para el metabolismo de varios aminoácidos importantes, es decir la metionina, cisteína, serina, glicina, e histidina. La síntesis de la metionina a partir de la homocisteína es catalizada por la metionina sintasa, una enzima que no solo el folato (como 5-metiltetrahidrofolato) pero también la vitamina B12. Por lo tanto, una deficiencia de folato (y/o vitamina B12) puede resultar en una disminución de la síntesis de metionina y una acumulación de homocisteína. Las concentraciones sanguíneas elevadas de homocisteína han sido consideradas por muchos años como un factor de riesgo para algunas enfermedades crónicas, incluyendo las enfermedades cardiovasculares y demencia (véase Prevención de Enfermedades).
Interacción con nutrientes
Vitamina B12 y vitamina B6
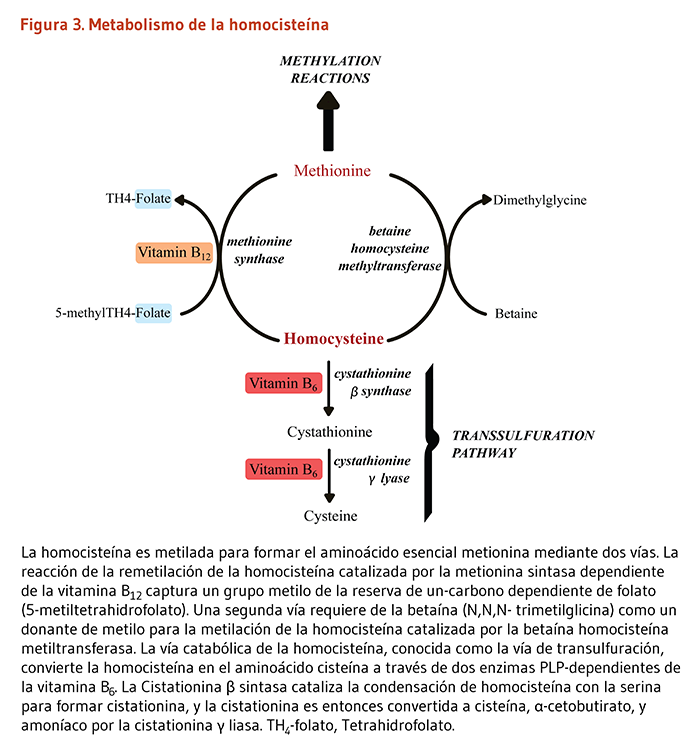
El metabolismo de la homocisteína, un intermediario en el metabolismo de los aminoácidos que contienen sulfuro, es un ejemplo de las interrelaciones entre los nutrientes necesarios para una salud y funcionalidad fisiológica óptimas. Los individuos sanos utilizan dos vías diferentes para metabolizar la homocisteína (Figura 3). Una vía (metionina sintasa) sintetiza la metionina de la homocisteína, y es dependiente tanto del folato como la vitamina B12 como cofactores. La otra vía convierte la homocisteína en otro aminoácido, denominado cisteína y requiere de dos enzimas dependientes de la vitamina B6. Por lo tanto, la cantidad de homocisteína en la sangre es regulada por tres vitaminas B: folato, vitamina B12, y vitamina B6 (4). En algunos individuos, la riboflavina (vitamina B2) también se relaciona en la regulación de las concentraciones de homocisteína (véase Riboflavina).
Riboflavina
Aunque es menos reconocida, el folato tiene una importante interacción metabólica con la riboflavina. La riboflavina es un precursor del flavín adenín dinucleótido (FAD), una coenzima necesaria para la actividad de las enzimas del folato encargadas de la metabolización, la 5,10-metilentetrahidrofolato reductasa (MTHFR). El FAD dependiente de la MTHFR en turno cataliza la reacción que genera el 5-metiltetrahidrofolato (ver Figura 2 arriba). Esta forma activa del folato es requerida para la formación de la metionina a partir de la homocisteína. Junto con otras vitaminas B, las ingestas más elevadas de riboflavina han sido asociadas con una disminución de las concentraciones de homocisteína en el plasma (5). Los efectos de la riboflavina en el metabolismo del folato parecen ser mayores en individuos homocigotos del polimorfismo común c.677C>T (genotipo TT) en el gen MTHFR (ver Variaciones genéticas en los requerimientos de folato) (6). Estos individuos (alrededor del 10% de los adultos a nivel mundial) típicamente presentan bajos estatus de folato, junto con elevadas concentraciones de homocisteína, particularmente cuando la ingesta de folato y/o riboflavina es subóptima. Las concentraciones elevadas de homocisteína en estos individuos, sin embargo, es altamente sensible a disminuir con la suplementación de riboflavina, confirmando la importancia de la interacción de la riboflavina y MTHFR (7).
Vitamina C
La vitamina C puede limitar la degradación de coenzimas folato naturales y ácido fólico suplementario en el estómago y así mejorar la biodisponibilidad del folato. Un ensayo cruzado en nueve hombres sanos encontró que la administración oral de ácido 5-metiltetrahidrofólico (343 μg) y vitamina C (289 mg o 974 mg) fue asociada con concentraciones más altas de folato en el suero en comparación con el ácido 5-metiltetrahidrofólico solo (8). Por otra parte, un reciente estudio sugirió que algunas variaciones genéticas del metabolismo del folato podrían influenciar el efecto de la vitamina C en el metabolismo del folato (9).
Biodisponibilidad
El folato dietario existe predominantemente en la forma de poliglutamilo (el cual contiene varios residuos de glutamato) mientras que el ácido fólico – la forma sintética de la vitamina- es un monoglutamato, es decir que contiene solo una fracción del glutamato. Además, folatos naturales son moléculas reducidas, mientras que el ácido fólico es completamente oxidado. Estas diferencias químicas tienen mayores implicaciones para la biodisponibilidad de la vitamina tal como que el ácido fólico es considerablemente más biodisponible que los folatos naturales en los alimentos a niveles de consumo equivalentes.
La absorción intestinal de folatos dietarios es un proceso de dos pasos que involucra la hidrolisis de poliglutamatos del folato a los derivados monoglutamil correspondientes, seguidos por su transporte dentro de las células intestinales. Allí, el ácido fólico es convertido en un folato natural, llamado 5-metiltetrahidrofolato, el cual es la mayor forma de folato circulante en el cuerpo humano (vea Figura 1 arriba).
La biodisponibilidad de folatos de procedencia natural es inherentemente limitada y variable. Existe mucha variabilidad en la facilidad con la que los folatos son liberados de diferentes matrices alimenticias, y la “cola” del poliglutamil es removida (desconjugación) antes de la absorción por las células intestinales. Además, otros componentes de la dieta pueden contribuir a la inestabilidad de los folatos lábiles durante los procesos de digestión. Como resultado, los folatos naturales muestran una biodisponibilidad incompleta en comparación con el ácido fólico. La biodisponibilidad del ácido fólico, en contraste, se asume es un 100% cuando este es ingerido como suplemento, mientras que el ácido fólico en alimentos fortificados se estima que posee alrededor de un 85% de la biodisponibilidad del ácido fólico suplementario.
Nótese que las recomendaciones del folato en los EE.UU. y otros ciertos países están ahora expresadas como Equivalentes de Folato Dietario (EFD), un cálculo que fue ideado para tener en cuenta la extensiva biodisponibilidad del ácido fólico en comparación con los folatos de origen natural (ver La Ingesta Diaria Recomendada).
Transporte
El folato y sus coenzimas requieren de transportadores para atravesar las membranas celulares. Los transportadores del folato incluyen el transportador de folato reducido (RFC), el transportador de folato acoplado a protones (PCFT), y proteínas receptoras del folato, FRα y FRβ. La homeostasis del folato es soportada por la distribución ubicua de los transportadores de folato, aunque la abundancia e importancia varía entre los tejidos (10). El PCFT juega un papel mayor en el transporte del folato intestinal debido a que las mutaciones que afectan el gen que codifica el PCFT causa una malabsorción hereditaria del folato. Además, un PCFT defectuoso conlleva a un deterioro en el transporte de folato al cerebro (ver Tratamiento de Enfermedades). La FRα y el RFC son también críticos para el transporte de folato a través de la barrera hematoencefálica cuando el folato extracelular es bajo o alto, respectivamente. El folato es esencial para el desarrollo adecuado del embrión y el feto. La placenta es conocida por concentrar el folato a la circulación fetal, llevando a concentraciones altas de folato en el feto a comparación a aquellas encontradas en la mujer embarazada. Los tres tipos de receptores han sido asociados con el transporte de folato a través de la placenta durante el embarazo (11).
Deficiencia
Causas
La mayoría de las veces, una deficiencia de folato es causada por una insuficiencia alimentaria; sin embargo, una deficiencia de folato puede también presentarse en un cierto número de otras situaciones. Por ejemplo, el consumo crónico y pesado de alcohol está asociado con una disminución de la absorción del folato (sumada a una ingesta dietaría baja), las cuales pueden conducir a una deficiencia de folato (12). Fumar esta también asociado con un estatus bajo de folato. En un estudio, las concentraciones de folato en la sangre fueron un 15% más bajas en fumadores que en no fumadores (13). Además, un transporte deteriorado del folato hacia el feto ha sido observado en mujeres embarazadas que fumaron o abusaron del alcohol durante el embarazo (14, 15).
El embarazo es el momento cuando él requerimiento de folato se incrementa en gran medida para mantener la demanda de la rápida replicación celular y para el crecimiento del feto, la placenta, y tejido materno. Ciertas condiciones como el cáncer o inflamaciones pueden derivar en un incremento en las tasas de división celular y metabolismo, causando un incremento en la demanda corporal por folato (16). Por otra parte, la deficiencia de folato puede resultar de condiciones de malabsorción, incluyendo enfermedades inflamatorias intestinales (enfermedad de Crohn y colitis ulcerosa) y la enfermedad celiaca (17). Varios medicamentos podrían también contribuir a una deficiencia de folato (véase Interacción con drogas). Finalmente, un cierto número de enfermedades genéticas que afectan la absorción, el transporte, o el metabolismo pueden causar una deficiencia de folato o impedir sus funciones metabólicas (véase Tratamiento de Enfermedades).
Síntomas
Una deficiencia clínica de folato conlleva a una anemia megaloblástica, la cual es reversible con un tratamiento con ácido fólico. Las células de división rápida como aquellas derivadas de la médula ósea son más vulnerables a los efectos de la deficiencia de folato debido a que la síntesis del ADN y la división celular dependen de las coenzimas del folato. Cuando el suministro de folato a las células de división rápida de la médula ósea es inadecuada, la división de glóbulos rojos es reducida, resultando en menos pero más grandes glóbulos rojos. Este tipo de anemia es llamada megaloblástica o anemia macrocítica, ya que hace referencia a glóbulos rojos inmaduros y agrandados. Los neutrófilos, un tipo de leucocito, se vuelven hipersegmentados, un cambio que pude ser observado al analizar microscópicamente una muestra de sangre. Debido a que los glóbulos rojos normales tienen un tiempo de vida en la circulación de aproximadamente cuatro meses, puede tomar meses para que los individuos deficientes de folato desarrollen la anemia megaloblástica característica. La progresión de este tipo de anemia conduce a una disminución de la capacidad de la sangre de transportar oxígeno y en última instancia, podría resultar en síntomas de fatiga, debilidad y dificultad para respirar (1). Es importante señalar que la anemia megaloblástica resultante de una deficiencia de folato es idéntica a la anemia megaloblástica resultante de una deficiencia de vitamina B12, y ensayos clínicos adicionales son requeridos para diagnosticar la verdadera causa de la anemia megaloblástica (véase Toxicidad).
Los individuos en etapas tempranas de una deficiencia de folato pueden no mostrar síntomas obvios, pero las concentraciones de homocisteína podrían incrementar (véase Prevención de Enfermedades). Sin embargo, la concentración de homocisteína circulante no es un indicador específico del estatus del folato, porque una homocisteína elevada puede ser el resultado de una deficiencia de vitamina B12 y de otras vitaminas B, factores del estilo de vida, e insuficiencia renal. Una deficiencia subclínica es comúnmente detectada midiendo las concentraciones de folato en el suero/plasma o en los glóbulos rojos.
La IDR
Determinación de la IDR
Tradicionalmente, el requerimiento de folato dietario fue definido como la cantidad necesaria para prevenir una deficiencia severa capaz de causar síntomas como la anemia. La ingesta diaria recomendada (IDR) más reciente (1998; ver Tabla 1) se basó principalmente en la adecuación de las concentraciones de folato presentes en glóbulos rojos a diferentes niveles de la ingesta de folato a juzgar por la ausencia de indicadores hematológicos anormales. Se ha demostrado que el folato de los glóbulos rojos se correlaciona con los depósitos hepáticos de folato y es usado como un indicador a largo plazo del estatus del folato. El folato en el plasma refleja la reciente ingesta de folato y no es un biomarcador seguro para el estatus del folato. El mantenimiento de niveles normales de homocisteína en la sangre, un indicador del metabolismo de un carbono, fue considerado sólo como un indicador auxiliar de la ingesta adecuada de folato.
Debido a que el embarazo es asociado con un aumento significativo en la división celular y otros procesos metabólicos que requieren coenzimas folato, la IDR para mujeres embarazadas es considerablemente más alta que para las mujeres que no están embarazadas (3). Sin embargo, la prevención de defectos del tubo neural no fue considerada cuando se estableció la IDR para mujeres embarazadas. Más bien la reducción del riesgo de defectos del tubo neural fue considerada una recomendación separada para mujeres capaces de quedar embarazadas (véase Prevención de Enfermedades), debido a que los eventos más importantes en el desarrollo del tubo neural ocurren antes de que muchas mujeres sean conscientes de su embarazo (18).
Equivalentes de Folato Dietario (EFD)
Cuando la Junta de Alimentos y Nutrición del Instituto de Medicina de los EE.UU. estableció la nueva recomendación dietaría de folato, ellos introdujeron una nueva unidad, el Equivalente de Folato Dietario (EFD) (1). El uso del EFD refleja una mayor biodisponibilidad del ácido fólico sintético encontrado en suplementos y alimentos fortificados, en comparación con la de folatos naturales presentes en los alimentos (18).
- 1 microgramo (μg) de folato alimentario aporta 1 μg de EFD
- 1 μg de ácido fólico ingerido con las comidas o como alimento fortificado aporta 1.7 μg de EFD
- 1 μg de ácido fólico (suplemento) ingerido con el estómago vacío aporta 2 μg de EFD
Por ejemplo una porción de alimento que contenga 60 μg de folato aportaría 60 μg de EFD, mientras una porción de tallarines fortificados con 60 μg de ácido fólico aportaría 1.7 x 60 = 102 μg de EFD, debido a una biodisponibilidad más alta de ácido fólico. Un suplemento de 400 μg de ácido fólico ingerido con el estómago vacío aportaría 800 μg de EFD. Debe tenerse en cuenta que los EFDs fueron determinados en estudios con adultos, y no se ha estudiado si el ácido fólico en la fórmula infantil es más biodisponible que los folatos en la leche materna. El uso de EFD para determinar el requerimiento de folato para infantes no sería deseable.
| Etapa de la Vida | Edad | Machos (μg/día) | Hembras (μg/día) |
|---|---|---|---|
| Infantes | 0-6 meses | 65 (IA) | 65 (IA) |
| Infantes | 7-12 meses | 80 (IA) | 80 (IA) |
| Niños | 1-3 años | 150 | 150 |
| Niños | 4-8 años | 200 | 200 |
| Niños | 9-13 años | 300 | 300 |
| Adolescentes | 14-18 años | 400 | 400 |
| Adultos | 19 años y más | 400 | 400 |
| Embarazo | Todas las edades | - | 600 |
| Período de lactancia | Todas las edades | - | 500 |
Variación genética en las requerimientos de folato
Un polimorfismo o variación común en la secuencia del gen para la enzima, 5,10-metilentetrahidrofolato reductasa (MTHFR), conocido como polimorfismo C677T MTHFR, resulta en una enzima termolábil (19). La sustitución de una citosina (C) por una tiamina (T) en el nucleótido 677 en el exón 4 del gen MTHFR lleva a una transición de alanina a valina en el dominio catalítico de la enzima. Dependiendo en la población, un 20% a 53% de los individuos podrían haber heredado una copia T (677C/T genotipo), y de un 3% a un 32% de los individuos podrían haber heredado dos copias T (677T/T genotipo) del gen MTHFR (20). La MTHFR cataliza la reducción de 5,10-5,10-metilentetrahidrofolato (5,10-metilen THF) en 5-metil tretrahidrofolato (5- MeTHF). Este último es la coenzima folato requerida para formar metionina de la homocisteína (ver Figura 2 arriba). La actividad de MTHFR es altamente disminuida en individuos heterocigóticos 677C/T (-30%) y homocigotos 677T/T (-65%) en comparación con aquellos con el genotipo 677C/C (21). La homocigocidad de la mutación (677T/T) está ligada a bajas concentraciones de folato en los eritrocitos y altas concentraciones de homocisteína (22, 23). El mejoramiento del estatus nutricional del folato en mujeres mayores con el alelo T redujo la concentración de homocisteína en el plasma (24). Una pregunta importante sin responder sobre el folato es si la presente IDR es suficiente para compensar por la actividad reducida de la enzima MTHR en individuos con al menos un alelo T, o si aquellos individuos tiene un requerimiento de folato superior que el IDR (25).
Prevención de Enfermedades
Complicaciones del embarazo
Defectos del tubo neural
El crecimiento y desarrollo fetal están caracterizados por una división celular generalizada. Disponer de suficiente folato es crítico para la síntesis del ADN y ARN. Los defectos del tubo neural (DTN) resultan del fracaso del cierre del tubo neural embrionario entre los días 21 y 28 después de la concepción, un tiempo en el que muchas mujeres no se dan cuenta de que están embarazadas (26). Los defectos del tubo neural incluyen varias malformaciones, como lesiones del cerebro (ej. anencefalia, encefalocele) o lesiones de la espina dorsal (espina bífida), los cuales son defectos de nacimiento devastadores, y algunas veces, fatales (27). La prevalencia de DTN en los EE.UU. previa a la fortificación de alimentos con ácido fólico se estimo fue 1 por cada 1,000 embarazos (1). Resultados de ensayos aleatorios han demostrado una reducción del 60% a 100% en casos de DTN cuando las mujeres consumieron suplementos de ácido fólico en adición a una dieta variada durante el período periconcepcional (aproximadamente un mes antes y al menos un mes después de la concepción) (28, 29). Los resultados de estos y otros estudios inspiraron al Servicio de Salud Pública de los EE.UU. a recomendar que todas las mujeres capaces de embarazarse consumieran 400 μg de ácido fólico diariamente para prevenir DTN. Las mujeres con un embarazo afectado previamente fueron también aconsejadas a recibir 4,000 μg de ácido fólico diariamente para así reducir la recurrencia de DTN (30). Estas recomendaciones fueron hechas para todas la mujeres en edad de procrear porque una cantidad folato adecuado debe estar disponible muy temprano en el embarazo, y porque muchos embarazos en los EE.UU. no son planeados (31).
A pesar de la efectividad de la suplementación con ácido fólico en el mejoramiento del estatus del folato, parece que globalmente solo un 30% de mujeres que se embarazan siguen correctamente la recomendación, y existe un poco de preocupación de que mujeres jóvenes de grupos étnicos minoritarios y niveles socioeconómicos más bajos son las menos propensas a seguir la recomendación (32-34). Para disminuir la incidencia de DTN, la legislación implementada por la FDA en 1998 exige la fortificación de todos los productos enriquecidos con granos con 1.4 mg de ácido fólico (véase Fuentes). El nivel de fortificación con ácido fólico requerido en los EE.UU. se estimó inicialmente que aportaba 100 μg de ácido fólico adicional a la dieta de una persona en promedio, aunque probablemente este aporte es aún mayor debido al abuso de ácido fólico por los fabricantes de alimentos (25, 35). La Red Nacional de la Prevención de Defectos del Nacimiento reporto aproximadamente una disminución del 30% en la frecuencia de DTN en los EE.UU. en comparación al periodo de pre-fortificación, y la prevalencia post-fortificación de DTN es 0.69 casos por cada 1,000 nacimientos vivos y muertes fetales (36).
También, un componente genético en la etiología de los DTN es evidenciado por el incremento del riesgo en mujeres con historial familiar de DTN y también por variaciones de riesgo entre etnias (37). Por otra parte, la ocurrencia de defectos del tubo neural puede ser atribuida a específicas interacciones gen-folato. El polimorfismo MTHFR c.677C>T y otras variaciones genéticas pueden incrementar el requerimiento de folato y susceptibilidad a un embarazo afectados con DTN. Previamente a la era de fortificación un estudio de caso y control mostro que ambas concentraciones de eritrocitos y folato del suero fueron significativamente más bajas en mujeres embarazadas con las variantes T/T y C/T en comparación al tipo natural del genotipo C/C (22), sugiriendo un metabolismo inadecuado del folato con genotipos maternos específicos. Un meta-análisis de 25 estudios de casos y controles, incluyendo 2,429 madres de caso y 3,570 madres de control, mostro una asociación positiva entre el polimorfismo maternal MTHFR c.677C>T y DTN (38). Otra variante del MTHFR, un cambio de A a C en la posición 1298, también ha sido asociado con una actividad reducida de MTHFR y un incremento en el riesgo de DTN (39). Los individuos heterocigotos para ambas de estas variantes (677C/T + 1298A/C) mostraron concentraciones más bajas de folato en el plasma y altas concentraciones de homocisteína que en los individuos con 677C/T + 1298A/A (40). Genotipos combinados con la homocigocidad G/G para el transportador de folato reducido (RFC-1) polimorfismo (c.80A>G) podrían contribuir aún más en la ocurrencia de DTN (41). El grado de riesgo de DTN fue también evaluado con polimorfismos MTHFR adicionales (c.116C>T, c.1793G>A) (42) como también con mutaciones que afectan otras enzimas del metabolismo de un carbono, incluyendo la metionina sintasa (MTR c.2756A>G) (43), metionina sintasa reductasa (MTRR c.66A>G) (44), y metilentetrahidrofolato deshidrogenasa (MTHFD1 c.1958G>A) (45). Mientras que el genotipo materno puede impactar el resultado del embarazo, parece que las interacciones de gen a gen entre la madre y el feto es influenciado aún más. El riesgo de DTN se incrementa debido a ciertas combinaciones genéticas, incluyendo interacciones materno (MTHFR c.677C>T)-fetal (MTHFR c.677C>T) y materno (MTRR c.66A>G)-fetal (MTHFR c.677C>T) (43, 44, 46). Finalmente, el estatus de la vitamina B12 ha sido asociado con la modificación del riesgo de DTN en la presencia de polimorfismos específicos en el metabolismo de un carbono (47).
Malformaciones cardiovasculares
Las anomalías congénitas del corazón son la mayor causa de mortalidad infantil pero también causan muertes en la adultez (48). Usando información de la base de datos del Registro Europeo de Anomalías Congénitas y Gemelos (EUROCAT), un estudio de caso y control, que involucro 596 casos y 2,359 controles, encontró que el consumo de por lo menos 400 μg/día de ácido fólico durante el periodo periconceptual (un mes antes de la concepción y durante 8 semanas de post-concepción, cubriendo el periodo embrionario del desarrollo del corazón) se asoció con un 18% en la disminución del riesgo de defectos congénitos del corazón (49). Recientes meta-análisis de 20 a 25 estudios de caso y control y basados en la familia observaron asociaciones positivas entre la variante MTHFR c.677C>T maternal, fetal, o paternal y en la incidencia de defectos congénitos del corazón (50, 51). Estudios adicionales son requeridos para elucidar los efectos de la interacción gen-nutriente en el riesgo de defectos congénitos del corazón; sin embargo, la investigación disponible actualmente indica que una ingesta adecuada de folato puede jugar un papel importante.
Fisuras orofaciales
El estatus materno de folato durante el embarazo puede influir en el riesgo de anomalías congénitas llamadas fisuras orofaciales, es decir fisura labial o labio leporino con o sin paladar hendido (52). Un estudio de caso y control poblacional en Noruega investigo el impacto de los suplementos de ácido fólico en madres de 377 recién nacidos con fisuras labiales con o sin paladar hendido, 196 con paladar hendido únicamente y 763 controles (53). Aunque ingestas dietarías o suplementos (durante los primero 3 meses de embarazo) por si solos no modifico significativamente el riesgo de fisuras labiales con o sin paladar hendido, el estudio reporto una riesgo 64% menor entre mujeres que tomaron multivitamínicos y suplementos de ácido fólico (≥400 μg diariamente) en adición a folatos dietarios. En la misma población, polimorfismos en el gen (c.699C>T) cistationina beta-sintasa (CBS) o el gen MTHFR (c677C>T; cuando la ingesta de folato estuvo bajo 400 μg/día) aparecieron protegidos, mientras que otras variantes del gen del metabolismo de un carbono del folato no pudieron ser ligados a las fisuras labiales con o sin paladar hendido (54, 55). Sin embargo, un reciente meta-análisis de 18 estudios mostro una elevación del riesgo de una fisura labial con o sin paladar hendido con la homocigocidad materna 677T/T (56). Estudios adicionales son necesarios para evaluar el riesgo de fisuras labiales con o sin paladar hendido integrando al mismo tiempo tanto el polimorfismo genético como los parámetros de la ingesta de folato. La evidencia epidemiológica que apoya el papel del folato en el riesgo de una fisura labial con paladar hendido es deficiente.
Otros resultados adversos del embarazo
Un bajo peso al nacer ha sido asociado con un incremento en el riesgo de mortalidad durante el primer año de vida y podría también influenciar los resultados en la salud en la edad adulta (57). Una reciente revisión sistemática y un meta-análisis de ocho ensayos controlados aleatorios encontraron una asociación positiva entre la suplementación de ácido fólico y el peso al nacer; ninguna asociación fue observada con respecto a la duración del periodo de gestación (58). Adicionalmente, un estudio prospectivo de cohorte de 306 adolescentes embarazadas asocio bajas ingestas de folato y el estatus del folato materno durante el tercer trimestre de embarazo con una alta incidencia de nacimientos pequeños para su edad gestacional (peso al nacer <10o percentil) (59). Por otra parte, el genotipo materno c677C>T MTHFR y un incremento en las concentraciones de homocisteína, considerados un indicador de la deficiencia de folato funcional, han sido ligados a bajos pesos al nacer (60).
Las concentraciones elevadas de homocisteína han sido también asociadas con un incremento en la incidencia de abortos espontáneos y otras complicaciones del embarazo, incluyendo preeclampsia y el desprendimiento de la placenta (61). Un exhaustivo estudio retrospectivo mostro que la homocisteína del plasma en mujeres Noruegas estuvo fuertemente relacionado a resultados y complicaciones adversas, incluyendo preeclampsia, parto prematuro, y un peso muy bajo al nacer, en embarazos previos (62). Un reciente meta-analisis de 51 estudios de cohorte prospectivos ligados a la variante c677C>T MTHFR con un riesgo incrementado de preeclampsia en poblaciones caucásicas y asiático orientales, reforzó la noción de que el metabolismo del folato puede jugar un papel en la condición (63). Un ensayo multicéntrico, controlado aleatorio, el Ensayo Clínico de Ácido Fólico (FACT) ha sido iniciado para evaluar si la suplementación diaria de hasta 5.1 mg de ácido fólico durante el embarazo podría prevenir preeclampsia y otro resultado adverso (ej. muerte materna, desprendimiento placentario, parto prematuro) en mujeres de alto riesgo (64). Una ingesta adecuada de folato durante el embarazo protege contra la anemia megaloblástica (65). Un reciente estudio de caso y control encontró una reducción en el riesgo de trastornos del espectro autista con un consumo de ácido fólico diario de 600 μg o más antes y durante el embarazo cuando la madre y el niño poseían el genotipo c677C>T del gen MTHFR (66).
De esta manera, es razonable el mantener la suplementación de ácido fólico a través del embarazo, incluso después de que el tubo neural haya cerrado con el fin de disminuir el riesgo de otros problemas durante el embarazo. Por otra parte, recientes revisiones sistemáticas de estudios basados en la observación no encontró evidencia alguna de una asociación entre la exposición al folato durante el embarazo y resultados adversos a la salud en el producto, en particular asma en la infancia y alergias (67, 68).
Enfermedades cardiovasculares
Homocisteína y enfermedades cardiovasculares
Los resultados de más de 80 estudios indicaron que incluso moderadas concentraciones de homocisteína en la sangre incrementan el riesgo de enfermedades cardiovasculares (ECV) (4). Posibles predisposiciones a accidentes vasculares han sido también ligadas a deficiencias genéticas en el metabolismo de la homocisteína en ciertas poblaciones (69). El mecanismo por el cual la homocisteína puede incrementar el riesgo de enfermedades vasculares ha sido el tema de una gran cantidad de investigación, pero podría involucrar efectos adversos de la homocisteína en la coagulación sanguínea, vasodilatación arterial, y engrosamiento de las paredes arteriales (70). Aunque un incremento en los niveles de homocisteína en la sangre se ha asociado consistentemente con el riesgo aumentado de enfermedades cardiovasculares, no está claro aún, si al disminuir los niveles de homocisteína se reducirá el riesgo de una enfermedad cardiovascular (véase Folato y homocisteína). Investigaciones han inicialmente predicho que un descenso prolongado en los niveles de homocisteína en el suero de 3 micromoles/litro reduciría el riesgo de ECV por hasta un 25% y sería un objetivo de tratamiento razonable para los individuos con alto riesgo (71, 72). Sin embargo, el análisis de ensayos clínicos recientes de la suplementación con vitaminas B ha mostrado que al disminuir las concentraciones de homocisteína no previno la ocurrencia de un segundo evento cardiovascular en pacientes con ECV existentes (73, 74). Consecuentemente, la Asociación Americana del Corazón recomienda la detección de niveles elevados de homocisteína total sólo en personas de "alto riesgo," por ejemplo, aquellos con historial personal o familiar de enfermedad cardiovascular prematura, malnutrición o síndromes de malabsorción, hipotiroidismo, falla renal, lupus, o personas que toman ciertos medicamentos (ácido nicotínico, teofilina, resinas fijadoras de ácidos biliares, metotrexato, y L-dopa).
Folato y homocisteína
Se ha asociado a las dietas ricas en folato con una disminución del riesgo de ECV, incluyendo la enfermedad coronaria cardíaca, infarto al miocardio (ataque al corazón), y accidente cerebrovascular. Un estudio que dio seguimiento a 1,980 hombres finlandeses por 10 años encontró que aquellos que consumieron más folato dietario tenían un riesgo 55% más bajo de un evento coronario agudo, en comparación con aquellos que consumieron menos folato dietario (75). De las tres vitaminas B que regulan las concentraciones de homocisteína, se ha demostrado que el ácido fólico tiene el mayor efecto en la disminución de los niveles basales de homocisteína en la sangre, cuando no hay una deficiencia de vitamina B12 o vitamina B6 coexistentes (véase Interacciones con nutrientes) (76). Se ha encontrado que un incremento en la ingesta de folato a través de alimentos ricos en folato o suplementos, reduce las concentraciones de homocisteína. Más aún, las concentraciones de homocisteína sanguínea han disminuido desde que la FDA ordenó la fortificación con ácido fólico del suministro de granos en los EE.UU. (25). Un meta-análisis de 25 ensayos controlados aleatorios, incluyendo casi 3,000 sujetos, encontró que la suplementación con 800 μg/día de ácido fólico podría alcanzar un máximo de un 25% en la disminución de las concentraciones de homocisteína en el plasma. En este meta-análisis dosis diarias de 200 μg y 400 μg de ácido fólico fueron asociadas con reducciones del 60% y 90% respectivamente, de homocisteína plasmática (78). Un régimen suplementario de 400 μg de ácido fólico, 2 mg de vitamina B6, y 6 μg de vitamina B12 ha sido ha sido recomendado por la Asociación Americana del Corazón, si una prueba inicial con una dieta rica en folato (véase Fuentes) no es exitosa en reducir adecuadamente las concentraciones de homocisteína (79).
Varios polimorfismos en el metabolismo de un carbono del folato modifican las concentraciones de homocisteína en la sangre (80). En particular el efecto de la variante c.677C>T de MTHFR ha sido examinada en relación a las políticas de fortificación del ácido fólico en todo el mundo. El análisis de ensayos aleatorios, incluyendo 59,995 sujetos sin un historial de ECV, revelo que la diferencia en las concentraciones de homocisteína entre los genotipos T/T y C/C fue mayor en regiones bajas en folato en comparación con regiones con políticas de fortificación de alimentos (3.12 vs. 0.13 micromoles/litro) (81). Aunque la suplementación con ácido fólico disminuye efectivamente las concentraciones de homocisteína, no está claro aún si también disminuye el riesgo de ECV. Un reciente meta-análisis de 19 ensayos clínicos aleatorios, incluyendo 47,921 sujetos con enfermedades cardiovasculares o renales preexistentes, encontró que la disminución de la homocisteína a través de la suplementación con ácido fólico y otra vitamina B fallo en reducir la incidencia de ECV a pesar de las significantes reducciones de las concentraciones de homocisteína en el plasma (74). Otros meta-análisis han confirmado la carencia de causalidad entre la disminución de homocisteína y el riesgo de ECV (80-82), incluyendo el riesgo de un accidente cerebrovascular (83, 84). Por consiguiente, La Asociación Americana del Corazón removió su recomendación del uso de ácido fólico para la prevención de enfermedades cardiovasculares en mujeres de alto riesgo (85). Debe notarse que la mayoría de ensayos preventivos a la fecha han sido realizados en pacientes con ECV con enfermedades avanzadas. La evidencia que apoya un papel beneficial del folato y relacionada con las vitaminas B parece ser más fuerte para la prevención primaria de accidentes cerebrovasculares (86). La introducción de la fortificación obligatoria con ácido fólico ha sido asociada con un decline en mortalidad relacionada a accidentes cerebrovasculares en Norte América, agregando más apoyo a el beneficio potencial del mejoramiento del estatus del folato y/o la disminución de homocisteína en la prevención de accidentes cerebrovasculares (87).
A pesar de la controversia sobre el papel de la disminución de la homocisteína en la prevención de ECV, algunos estudios han investigado el efecto de la suplementación del ácido fólico en el desarrollo de la aterosclerosis, un conocido factor de riesgo en accidentes vasculares. La medición del grosor íntima-media carotídeo (GIMc) es un objetivo subrogado para la aterosclerosis temprana y un pronosticador para eventos cardiovasculares (88). El meta-análisis de 10 ensayos aleatorios que probaron el efecto de la suplementación con ácido fólico mostro una significante reducción del GIMc en sujetos con enfermedades renales crónicas y en aquellos en riesgo de padecer de EC, pero no en participantes saludables (89). La disfunción endotelial es una característica común en la aterosclerosis y enfermedad vascular. Altas dosis de ácido fólico (400-10,000 μg/día) han sido asociadas con mejoramientos de la salud vascular en sujetos sanos y con ECV (90). Aunque recientes ensayos fallaron en demostrar alguna protección de enfermedades cardiovasculares de la suplementación con ácido fólico, una ingesta baja de folato es un conocido factor de riesgo para las enfermedades cardiovasculares y más investigación es necesaria para explorar el papel del folato en el mantenimiento de la salud vascular (91).
Cáncer
Se piensa que el cáncer surge de un daño al ADN por el exceso en la reparación continua de éste y/o de la expresión inadecuada de genes fundamentales. Debido a las importantes funciones que desempeña el folato en la síntesis y metilación del ADN y ARN, es posible que la ingesta inadecuada de folato contribuya a la inestabilidad del genoma y rotura de cromosomas que a menudo caracteriza el desarrollo del cáncer. En particular, la replicación y reparación del ADN son críticas para el mantenimiento del genoma, y la escasez en los nucleótidos causada por la deficiencia de folato podría llevar a una inestabilidad del genoma y mutaciones en el ADN. Una disminución de 5,10-metileno THF puede comprometer la conversión de desoxiuridina monofosfato (dUMP) a desoxitimidina monofosfato (dTMP) por la enzima timidilato sintasa (TS), causando una acumulación de uracilo y una reducción de timina. Esto podría entonces llevar a una incorporación errónea en el ADN durante la replicación o reparación, y causar un daño al ADN, incluyendo mutaciones puntuales y roturas de la cadena (92). Debido a que la 5,10-metileno THF es también el sustrato de la enzima MTHFR, es plausible que una reducción de la actividad de MTHFR con el polimorfismo c.677C>T pudiese incrementar el uso de 5,10-metileno THF para la síntesis de timidilato y prevención de daño al ADN. Sin embargo, la hipótesis podría solo ser validad en una situación de deficiencia de folato (93). Al contrario, se argumentó que la suplementación con ácido fólico podría alimentar la síntesis de ADN, de este modo promoviendo el crecimiento tumoral. Esto es apoyado por la observación de que la TS puede funcionar como un promotor de tumores (oncogén), mientras que una reducción en la actividad de la TS está ligada a un menor riesgo de cáncer (94, 95). Adicionalmente, las moléculas anti-folato que bloquean la vía de la síntesis de timidilato se utilizan con éxito en la terapia contra el cáncer (96). El folato también controla el ciclo de la homocisteína/metionina y el depósito de S-adenosilmetionina (SAM), el donante de metilo para las reacciones de metilación. De esta manera, la deficiencia de folato puede perjudicar la metilación del ADN y proteínas y alterar la expresión de genes involucrados en la reparación del ADN, proliferación y muerte celular. La hipometilación global del ADN, una característica distintiva del cáncer, causa inestabilidad del genoma y ruptura de cromosomas (revisado en 97).
El consumo de por lo menos cinco porciones de frutas y vegetales diariamente ha sido consistentemente asociado con una disminución de la incidencia de cáncer (98). Las frutas y vegetales son excelentes fuentes de folato, las cuales pueden jugar un papel en su efecto anti-cancerígeno. Estudios basados en la observación han encontrado que un estatus disminuido del folato está asociado con canceres de sitio específico. Mientras que la fortificación de alimentos es obligatoria en los EE.UU. (desde 1998; vea Fuentes), preocupaciones acerca del impacto de altas ingestas de ácido fólico en la salud han retrasado la practica en otros países (99). Sin embargo, los meta-análisis más recientes de ensayos de intervención del ácido fólico (dosis suplementarias de entre 500 a 5,000 μg/día durante al menos un año) no mostro algún beneficio especifico o daño con respecto a la incidencia de cáncer total o de sitio especifico (100, 101).
Cáncer colorrectal
Un análisis combinado de 13 estudios de cohorte prospectivos, los cuales dieron seguimiento a un total de 725,134 individuos por un periodo de 7 a 20 años, revelo una modesta asociación inversa entre la ingesta dietaría y total (proveniente de alimentos y suplementos) de folato y el riesgo de cáncer de colon. Específicamente, se estimó un decline del 2% del riesgo de cáncer de colon por cada incremento de 100 μg/día en la ingesta total de folato (102). Un exhaustivo estudio prospectivo estadounidense, el cual dio seguimiento a 525,488 sujetos, de edades entre 50 a 71 años entre 1995 y 2006, correlacionaron el folato dietario, ácido fólico suplementario, e ingestas totales de folato con una disminución del riesgo de cáncer colorrectal (103). Sin embargo, cuando se estratifico por género, no hubo asociación entre la ingesta de folato dietario y el riesgo de cáncer colorrectal en mujeres (103, 104). Una falta de asociación entre el riesgo de cáncer colorrectal e ingestas dietarías, suplementarias, y totales de folato, fue también reportada en otro estudio perspectivo que dio seguimiento a más de 90,000 mujeres postmenopáusicas estadounidenses durante un periodo de 11 años abarcando los periodos de pre y post fortificación (105). Estos datos sugirieron la posible influencia de género sobre la modificación del riesgo de cáncer colorrectal por el folato. En el último estudio, una elevación significante pero transitoria del riesgo fue también observada durante la era de post-fortificación; sin embargo algunos han afirmado que es poco probable que esto sea causado por una ingesta incrementada de folato debido a la fortificación obligatoria (106). Finalmente, un meta-análisis de 18 estudios de casos de controles encontró una ligera reducción en el riesgo de cáncer colorrectal con folato proveniente de alimentos (107). Sin embargo, es importante tener en cuenta que los estudios de caso y control fueron altamente heterogéneos, y que los autores declararon que la fibra dietaría, vitaminas, y el consumo de alcohol pudieran haber alterado sus resultados. Por otra parte, el límite más bajo del cuantil más alto de la ingesta de folato fue altamente variable, oscilando de entre 270 a 1,367 μg/día (107).
Mientras que la mayoría de la investigación epidemiológica muestra un efecto protector del folato en contra del desarrollo del cáncer colorrectal, se ha sugerido que las altas dosis de ácido fólico suplementario en realidad podrían acelerar el crecimiento del tumor en pacientes con cáncer (108). Mientras que un estatus más alto de folato dentro el rango dietario normal es extensamente considerado como protector contra el cáncer, algunos investigadores permanecen preocupados de que la exposición excesiva de altas ingestas de ácido fólico podrían incrementar el crecimiento de neoplasias pre-existentes (108). Varios ensayos clínicos abordaron el efecto de la suplementación de ácido fólico en pacientes con un historial de adenoma colorrectal, con ensayos que encontraron una reducción del riesgo o ningún efecto del ácido fólico suplementario (109-112). Un reciente meta-análisis de tres ensayos controlados aleatorios exhaustivos en sujetos de alto riesgo no demostró algún incremento en la recurrencia de adenoma colorrectal en sujetos suplementados con 500 o 1,000 μg/día de ácido fólico por 24 a 42 meses cuando se comparó con el tratamiento con placebo (113).
Como se sugirió anteriormente, el genotipo MTHFR 677T/T podría prevenir la incorporación errónea de uracilo y proteger la integridad y estabilidad del ADN bajo condiciones de folato bajo. Un meta-análisis de 62 estudios de caso-control y dos de cohorte revelaron que mientras la variante T/T reduce el riesgo de cáncer colorrectal un 12% en comparación a ambos C/T y C/C genotipos, el riesgo se redujo un 30% con altas (348-1,583 μg/día) frente a ingestas bajas totales de folato (264-450 μg/día), independientemente del genotipo (114). Un polimorfismo común (c.2756A>G) en el gen MTR, el cual codifica para la metionina sintasa, fue también examinado en relación con el riesgo de adenoma colorrectal y cáncer. La metionina sintasa cataliza la conversión simultánea de homocisteína y 5-metileno THF en metionina y TFH, respectivamente. El reciente meta-análisis de 27 estudios de caso y control mostro ninguna asociación entre la variante del MTR y el riesgo de cáncer (115).
Aunque el consumo de alcohol interfiere con la absorción y el metabolismo del folato (16), un estudio de caso y control y cinco estudios de cohorte prospectivos han reportado o una reducción en el riesgo de cáncer colorrectal entre no bebedores en comparación a bebedores o una falta de asociación (107). Sin embargo, un exhaustivo estudio prospectivo que dio seguimiento a más de 28,000 profesionales de la salud masculinos por 22 años, una ingesta de más de dos bebidas alcohólicas (>30 gramos de alcohol) por día aumento el riesgo de cáncer colorrectal por un 42% durante el periodo de pre-fortificación. El riesgo de cáncer colorrectal no se incrementó durante el periodo de post-fortificación, sugiriendo que es la combinación de una ingesta alta de alcohol y baja en folato que podría incrementar el riesgo de cáncer colorrectal. A pesar de todo, otro estudio perspectivo que dio seguimiento a más de 69,000 enfermeras por 28 años no reporto un incremento significante del riesgo de cáncer colorrectal con una ingesta de alcohol antes y después de la fortificación con ácido fólico obligatoria (116). En algunos estudios, individuos que son homocigotos para el polimorfismo c.677C>T MTHFR (T/T) se ha encontrado que están a un riesgo disminuido de padecer cáncer colorrectal cuando la ingesta de folato es adecuada. Sin embargo se ha descubierto que cuando la ingesta de folato es baja o la ingesta de alcohol es alta, los individuos con el genotipo (T/T) se encuentran en un riesgo incrementado de cáncer colorrectal (117, 118).
Cáncer de seno
Varios estudios de cohorte prospectivos y de caso y control han reportado resultados mixtos al investigar si la ingesta de folato afecta el riesgo de cáncer de seno (119). Un meta-análisis de 15 estudios prospectivos y un estudio caso-control anidado no encontró relación alguna con la ingesta de folato dietaría (120). Una ingesta de alcohol moderada ha sido asociada con un incremento en el riesgo de cáncer de seno en mujeres (121). Los resultados de tres estudios prospectivos sugirieron que una ingesta incrementada de folato puede reducir el riesgo de cáncer de seno en mujeres que regularmente consumen alcohol (122-124). De esta manera, una ingesta alta de folato podría estar asociada con una reducción del riesgo solo en mujeres cuyo riesgo de cáncer es incrementado por el consumo de alcohol. Un estudio prospectivo bastante exhaustivo en más de 88,000 enfermeras reporto que una ingesta de ácido fólico no estaba asociada con cáncer de seno en mujeres que consumieron menos de una bebida alcohólica por día. Sin embargo, en mujeres que consumieron por lo menos una bebida alcohólica por día, la ingesta de ácido fólico de al menos 600 μg diariamente resulto en aproximadamente la mitad del riesgo de cáncer de seno en comparación con las mujeres que consumieron 300 μg de ácido fólico diariamente (124). No obstante, si y como el consumo de alcohol incrementa el riesgo de cáncer de seno es aún un tema de discusión (125, 126). Finalmente, recientes meta-análisis que evaluaron la influencia de polimorfismos en el metabolismo de un carbono en el riesgo de cáncer de seno encontraron que variantes específicas en el gen que codifica la timidilato sintasa incrementaron el riesgo de cáncer de seno en ciertas poblaciones étnicas (127, 128).
Cáncer infantil
La incidencia de tumores de Wilm (cáncer de riñón) y ciertos tipos de cánceres cerebrales (neuroblastoma, ganglioneuroblastoma, y ependimoma) en niños ha disminuido desde la fortificación obligatoria del suministro de granos estadounidense en 1998 (129). Sin embargo, las tasas de incidencia se mantuvieron sin cambios entre los periodos de pre y post fortificación para la leucemia- una malignidad predominante en la infancia. A pesar de estudios tempranos que ligaban la suplementación de ácido fólico materna durante el embarazo con la disminución del riesgo de leucemia infantil, investigaciones más recientes han encontrado poca evidencia para apoyar un efecto preventivo del ácido fólico (130). Varios meta-análisis también han encontrado poco a ningún efecto protector con los polimorfismos de MTHFR; sin embargo el meta-análisis más reciente de 22 estudios de caso y control encontró una reducción en el riesgo de leucemia linfoblástica aguda (LLA) con la variante c.677C>T en caucásicos y asiáticos (131).
Enfermedad de Alzheimer y deterioro cognitivo
La enfermedad de Alzheimer es la forma de demencia más común, afectando más de 5 millones de individuos mayores de 65 años de edad en los EE.UU. (132). Se han asociado la deposición de la placa beta-amiloide, la proteína formadora de ovillos Tau, y un incremento de la muerte celular en el cerebro de los pacientes con Alzheimer con el descenso cognitivo y la perdida de la memoria. Un estudio asocio el incremento del consumo de frutas y verduras, las cuales son fuentes abundantes de folato con un riesgo reducido de desarrollar demencia y Alzheimer en mujeres (133). A través de su papel en la síntesis de ácidos nucleicos y la provisión de donantes de metilo para las reacciones de metilación, el folato es crítico para el funcionamiento y desarrollo normal del cerebro, no solamente durante el embarazo y después del nacimiento, sino también posteriormente en la vida (134). En un estudio de corte transversal en mujeres de edad avanzada, los pacientes con Alzheimer tuvieron concentraciones significantemente más altas de homocisteína y concentraciones más bajas de folato en los glóbulos rojos en comparación con individuos saludables. Sin embargo, no hubo diferencia en el nivel de folato en el suero entre los grupos, sugiriendo que el estatus de folato a largo plazo en lugar de la ingesta reciente de folato, podría estar asociado con el riesgo de Alzheimer (135).
Varios investigadores han descrito asociaciones entre el incremento de las concentraciones de homocisteína y el deterioro cognitivo en las personas mayores (136), pero estudios de cohorte prospectivos no han encontrado que altas ingestas de folato estén asociadas con un mejoramiento de la cognición (137, 138). Altas concentraciones de homocisteína fueron encontradas en individuos que sufrían de demencia, incluyendo Alzheimer, y demencia vascular, en comparación a sujetos sanos (139, 140). Aunque las deficiencias de folato, vitamina B12, y vitamina B6 podrían incrementar las concentraciones de homocisteína, una reducción en las concentraciones de vitamina en el suero de pacientes con Alzheimer en comparación a individuos sanos no pudo ser atribuido a una disminución de las ingestas de vitaminas (141). No está claro actualmente si la homocisteína sérica es un factor de riesgo para el desarrollo de la demencia o simplemente está asociada con la disminución cognitiva. En la última década, un cierto número de ensayos clínicos han probado el uso de las vitaminas B para disminuir la homocisteína y prevenir o retrasar el deterioro cognitivo. Un meta-análisis de nueve ensayos controlados aleatorios con placebo de la suplementación con ácido fólico (0.2 a 15 mg/día por una duración media de seis meses) en individuos mayores de 45 años de edad fallo en encontrar un efecto a corto plazo en las funciones cognitivas, incluyendo memoria, velocidad, lenguaje, y funciones ejecutivas (142). Más recientemente, un meta-análisis de 19 ensayos controlados aleatorios con placebo de la suplementación con vitaminas B no encontró alguna diferencia en los parámetros cognitivos entre los grupos de tratamiento y placebo, a pesar de que el tratamiento efectivamente disminuyo las concentraciones de homocisteína (143). Hallazgos inconsistentes a través de los ensayos puede deberse a diferencias en el diseño y metodología (revisado en 144).
No obstante, un ensayo controlado aleatorio con placebo de dos años en 168 sujetos de edad avanzada con deterioro cognitivo leve describió los beneficios de un régimen diario de 800 μg de ácido fólico, 500 μg de vitamina B12 y 20 mg de vitamina B6 (145, 146). La atrofia de regiones cerebrales especificas afectadas por el Alzheimer fue observada en individuos de ambos grupos, y esta atrofia estaba correlacionada con el deterioro cognitivo; sin embargo, el grupo de tratamiento con vitaminas-B experimento una menor perdida de materia gris en comparación a el grupo de placebo (0.5% vs. 3.7%). Un beneficio mayor fue observado en sujetos con concentraciones mayores de homocisteína al inicio del estudio, sugiriendo la importancia de la disminución de homocisteína circulante en la prevención del deterioro cognitivo y la demencia. Aunque alentador, el efecto de la suplementación con vitaminas B necesita ser estudiado más a fondo en ensayos exhaustivos que evalúen resultados a largo plazo, como la incidencia del Alzheimer.
Tratamiento de Enfermedades
Enfermedades metabólicas
El ácido folínico (vea Figura 1 arriba), un derivado del ácido tetrahidrofólico, es usado en el manejo clínico de errores congénitos raros que afectan el transporte y metabolismo del folato (revisado en 147). Tales condiciones son de una herencia autosómica recesiva, lo que significa que solo individuos que reciben dos copias del gen mutado (uno de cada padre) desarrollan la enfermedad.
Malabsorción del folato hereditaria
La malabsorción del folato hereditaria es causada por mutaciones en la codificación del gen SLC46A1 para el trasportador de folato PCFT y típicamente afecta la absorción gastrointestinal del folato y el transporte del folato en el cerebro (148). Los pacientes se presentan con bajas o indetectables concentraciones de folato en el suero y en el fluido cerebroespinal, pancitopenia (baja cantidad de todos los glóbulos rojos), respuestas inmunes deterioradas que incrementan la susceptibilidad a infecciones, y un fallo general para prosperar (149). Síntomas neurológicos incluyendo convulsiones, han sido también observadas (150). Mejoras clínicas han sido registradas tras la suministración parenteral de ácido folínico (151).
Síndrome de la deficiencia de folato cerebral (DFC)
La DFC se caracteriza por bajos niveles de coenzimas de folato en el fluido cerebroespinal a pesar de concentraciones normales de folato en la sangre. El transporte de folato a través de la barrera hematoencefálica está comprometida en la DFC y ha sido ligada ya sea a la presencia de anticuerpos bloqueando el receptor de folato FRα o a mutaciones en el gen FOLR1 que codifica el FRα (152, 153). Anormalidades neurológicas, junto con deficiencias visuales y auditivas, han sido descritas en niños con DFC; el desorden del espectro autista (TEA) está también presente en algunos casos. El ácido folínico (también conocido como leucovorina) puede entrar en el cerebro y normalizar el nivel de coenzimas de folato y se ha mostrado que normaliza las concentraciones de folato y mejora varias interacciones sociales en la DFC, incluyendo el estado de ánimo, comportamiento, y comunicación verbal en niños con (TEA) (152, 154, 155).
Deficiencia de dihidrofolato reductasa (DHFR)
La DHFR es la enzima dependiente de la NADPH que cataliza la reducción del ácido dihidrofólico (DHF) a ácido tetrahidrofólico (THF). La DHFR es también requerida para convertir el ácido fólico a DHF. La deficiencia de DHFR está caracterizada por una anemia megaloblástica y una deficiencia de folato cerebral causando convulsiones intratables y déficits mentales. Aunque el tratamiento con ácido folínico puede aliviar los síntomas de la deficiencia de DHFR, un diagnóstico temprano es esencial para prevenir un daño irreversible al cerebro y mejorar los resultados clínicos (156, 157).
Fuentes
Fuentes alimenticias
Las verduras de hojas verdes (follaje) son fuentes ricas en folato y proporcionan la base para su nombre. Los jugos de frutas cítricas, las legumbres, y los alimentos fortificados son también excelentes fuentes de folato (1); el contenido de folato de cereales fortificados varía en gran medida. La siguiente tabla muestra una serie de alimentos ricos en folato, junto con su contenido de folato en microgramos (μg; La siguiente tabla muestra ). Para mayor información del contenido de nutrientes de alimentos específicos, busque en la base de datos de composición de los alimentos del USDA.
| Alimento | Porción | Folato (μg) |
|---|---|---|
| Lentejas (semillas maduras, cocidas, hervidas) | ½ taza | 179 |
| Garbanzos (cocidos, hervidos) | ½ taza | 141 |
| Espárragos (cocidos, hervidos) | ½ taza (~6 varitas) | 134 |
| Espinacas (cocidas, hervidas) | ½ taza | 131 |
| Habas (grandes, semillas maduras, cocidas, hervidas) | ½ taza | 78 |
| Jugo de naranja (recién extraído) | 6 onzas | 56 |
| Espagueti (enriquecido, cocido) | 1 taza | 167* |
| Arroz blanco (enriquecido, cocido) | 1 taza | 153* |
| Pan (enriquecido) | 1 rebana | 84* |
| *Para ayudar a prevenir defectos del tubo neural, la FDA ordenó que se agregara 1.4 miligramos (mg) de ácido fólico por cada kilogramos (kg) de cereal a los productos de grano refinado, los que ya estaban enriquecidos con niacina, tiamina, riboflavina, y hierro desde el 1ro de enero de 1998. La adición de nutrientes en los alimentos con el fin de prevenir una deficiencia nutricional, o restaurar los nutrientes perdidos en el proceso, se conoce como fortificación. La FDA inicialmente estimo que este nivel de fortificación incrementaría la ingesta dietaría 100 μg de ácido fólico al día en promedio. Sin embargo evaluaciones adicionales basadas en estudios basados en la observación sugirieron aumentos del doble que lo predicho por la FDA (35). La prevalencia de bajas concentraciones de folato en ambos suero y eritrocitos es actualmente menos de 1% en la población de los EE.UU., en comparación a 24% y 3.5% respectivamente antes de los periodos de fortificación (158). | ||
Suplementos
La principal forma de folato suplementario es el ácido fólico. Este se encuentra disponible en un sólo ingrediente y en productos combinados como vitaminas del complejo B y multivitamínicos. Dosis iguales o superiores a 1 mg requieren de receta médica (159). Adicionalmente, el ácido folínico, un derivado del ácido tetrahidrofólico, es usado para el manejo de ciertas enfermedades metabólicas (véase Tratamiento de Enfermedades). Ademas, la FDA estadounidense ha aprobado la suplementación de folato en anticonceptivos orales. La adición de levomefolato de calcio (la sal de calcio del meTHF; 451 μg/tableta) a anticonceptivos orales está destinada a elevar el nivel de folato en las mujeres en edad fértil (160). De acuerdo a un estudio nacional estadounidense, solo un 24% de mujeres no embarazadas entre la edad de 15-44 años están cumpliendo con la recomendación actual de 400 μg/día de ácido fólico (161).
Seguridad
Toxicidad
No hay efectos adversos asociados con el consumo en exceso de folato proveniente de alimentos. Las preocupaciones en cuanto a su seguridad se limitan a la ingesta de ácido fólico sintético. Una deficiencia de vitamina B12, aunque con frecuencia sin diagnosticar, podría afectar a un número significativo de personas, especialmente a adultos mayores (véase el artículo separado en Vitamina B12). Uno de los síntomas de la deficiencia de vitamina B12 es la anemia megaloblástica, la cual es indistinguible de la que se asocia con la deficiencia de folato (véase Deficiencia). Grandes dosis de ácido fólico, administradas a un individuo con deficiencia de vitamina B12 no diagnosticada, pueden corregir la anemia megaloblástica sin corregir la deficiencia de vitamina B12 subyacente, dejando al individuo en riesgo de desarrollar un daño neurológico irreversible. Tales casos de progresión neurológica en la deficiencia de vitamina B12, se han visto en su mayoría en dosis de ácido fólico de 5,000 μg (5 mg) o más altas. Con el fin de asegurar la prevención de un daño neurológico irreversible en personas con deficiencia de vitamina B12, La Junta de Nutrición y Alimentos del Instituto de Medicina estadounidense recomienda que todos los adultos limiten su ingesta de ácido fólico (suplementos y fortificación) a 1,000 μg (1 mg diario; Tabla 3). La junta también señaló que la deficiencia de vitamina B12, es muy rara en mujeres en edad fértil, haciendo poco probable que un consumo de ácido fólico igual o mayor a 1,000 μg/día, cause problemas (1); sin embargo, la información sobre los efectos de grandes dosis es limitada.
| Grupo Etario | NM (μg/día) |
|---|---|
| Infantes 0-12 meses | No es posible establecer* |
| Niños 1-3 años | 300 |
| Niños 4-8 años | 400 |
| Niños 9-13 años | 600 |
| Adolescentes 14-18 años | 800 |
| Adultos 19 años o más | 1,000 |
| *La fuente de ingesta debiese ser solo de alimentos y formula. | |
La saturación de la capacidad metabólica de la DHFR por dosis orales de ácido fólico ha sido asociada con la aparición de ácido fólico sin metabolizar en la sangre (162). Alteraciones hematológicas y una cognición más pobre han sido asociadas con la presencia de ácido fólico sin metabolizar en adultos mayores (≥60 años) con una deficiencia de vitamina B12 (163, 164). Un estudio pequeño conducido en mujeres postmenopáusicas también planteo preocupaciones acerca del efecto de la exposición de ácido fólico sin metabolizar en la función inmune (165). En un pequeño ensayo aleatorio abierto en 38 mujeres en edad reproductiva que recibieron 30 semanas de suplementos multivitamínicos diarios, una suplementación diaria con ya sea 1.1 mg o 5mg de ácido fólico resulto en la aparición transitoria de ácido fólico sin metabolizar en la sangre en las primeras 12 semanas de la suplementación (166). Sin embargo las concentraciones de ácido fólico sin metabolizar regresaron a los niveles del inicio del estudio al final del estudio, sugiriendo que los mecanismos de adaptación eventualmente convirtieron el ácido fólico a formas reducidas de folato. No obstante, el uso de levomefolato (5-metil THF) suplementario podría proveer de una alternativa para prevenir los potenciales efectos negativos del ácido fólico sin convertir en adultos mayores.
Interacción con drogas/fármacos
Cuando se toman drogas anti-inflamatorias no esteroidales (AINEs), como la aspirina o el ibuprofeno, en dosis terapéuticas muy grandes (por ejemplo, para tratar artritis severa), éstas podrían interferir con el metabolismo del folato. En comparación, no se ha demostrado que el uso rutinario de AINEs en bajas dosis afecte negativamente el estatus del folato. Se ha demostrado que el anticonvulsivo fenitoina inhibe la absorción intestinal de folato, y varios estudios han asociado un estado de folato disminuido con el uso a largo plazo de los anticonvulsivos fenitoina, fenobarbital, y pirimidona (167). Sin embargo, pocos estudios controlaron las diferencias en la ingesta de folato dietario, entre los usuarios, y los no usuarios de anticonvulsivos. Además, tomar ácido fólico al mismo tiempo que los agentes para reducir colesterol, colestiramina y colestipol, podría disminuir la absorción de ácido fólico (159). El metotrexato es un antagonista del ácido fólico, usado para tratar una serie de enfermedades incluyendo cáncer, artritis reumatoide y psoriasis. Algunos de los efectos secundarios del metotrexato son parecidos a los de una deficiencia de folato severa, y la suplementación con ácido fólico o folínico es usada para reducir la toxicidad del anti-folato. Otras moléculas anti-folato actualmente usadas en la terapia contra el cáncer incluyen aminopterina, pemetrexed, y raltitrexed (96). Además se ha demostrado que una serie de otros medicamentos tienen una actividad anti-folato, incluyendo trimetoprima (un antibiótico), pirimetamina (un antimalarial), triamtereno (un medicamento para la presión sanguínea), y sulfasalazina (un tratamiento para la colitis ulcerosa). Los primeros estudios de anticonceptivos orales (píldoras anticonceptivas), que contenían altas dosis de estrógeno, mostraron efectos adversos sobre los niveles del folato; sin embargo, estos hallazgos no han sido respaldados por estudios más recientes con bajas dosis de anticonceptivos donde la ingesta de folato fue controlada (168).
Recomendación del LPI
La evidencia científica disponible demuestra que una ingesta de folato adecuada previene defectos del tubo neural y otros problemas del embarazo, es útil en la disminución del riesgo de algunas formas de cáncer, especialmente en personas genéticamente susceptibles, y que podría reducir el riesgo de enfermedades cardiovasculares. El Instituto Linus Pauling (LPI) recomienda que los adultos tomen un suplemento multivitamínico-mineral diario, el cual típicamente contiene 400 μg de ácido fólico, el Valor Diario (VD). Incluso con una ingesta de ácido fólico mayor al promedio de alimentos fortificados, es poco probable que la ingesta de ácido fólico diaria de una persona regularmente exceda el nivel de ingesta máxima de 1,000 μg/día, establecida por la Junta de Nutrición y Alimentos (véase Seguridad).
Adultos mayores (>50 años)
La recomendación de 400 μg/día de ácido fólico suplementario, como parte de un suplemento multivitamínico-mineral diario, además de una dieta rica en folato, es especialmente importante en adultos mayores debido a que los niveles de homocisteína sanguínea tienden a aumentar con la edad (véase Prevención de Enfermedades).
Autores y Críticos
Escrito en 2000 por:
Jane Higdon, Ph.D.
Instituto Linus Pauling
Universidad Estatal de Oregon
Actualizado en Abril de 2002 por:
Jane Higdon, Ph.D.
Instituto Linus Pauling
Universidad Estatal de Oregon
Actualizado en Septiembre de 2007 por:
Victoria J. Drake, Ph.D.
Instituto Linus Pauling
Universidad Estatal de Oregon
Actualizado en Junio de 2014 por:
Barbara Delage, Ph.D.
Instituto Linus Pauling
Universidad Estatal de Oregon
Revisado en Diciembre de 2014 por:
Helene McNulty, Ph.D., R.D.
Profesor de Nutrición Humana y Dietética
Centro para la Alimentación y Salud de Irlanda del Norte (NICHE)
Universidad de Ulster
Coleraine, Reino Unido
Traducido al Español en 2015 por:
Silvia Vazquez Lima
Instituto Linus Pauling
Universidad Estatal de Oregon
La actualización del 2014 de este artículo fue suscrito, por una subvención de Bayer Consumer Care AG, Basel, Suiza.
Originalmente traducido al español en 2012 por Guillermo Sandoval y editado por Andrew Quest (Ph.D.) y Lisette Leyton (Ph.D.), todos provenientes de la Universidad de Chile. Estos esfuerzos fueron patrocinados por el projecto Anillo #ACT1111, CONICYT-Chile, programa PIA.
Derechos de autoría 2009-2026 Instituto Linus Pauling
Figure Alternative Text
Figure 1. Chemical structures
The figure shows chemical structures of three forms of folate, including synthetic compounds: folic acid (C19H19N7O6), 5-methyltetrahydrofolate (C20H25N7O6), and folinic acid (C20H23N7O7).
Figure 2. Overview of one-carbon metabolism
The figure illustrates a biochemical pathway of one-carbon metabolism. At the top of the figure, folate is reduced to dihydrofolate and then further reduced to tetrahydrofolate, which enters a circular pathway. Through a vitamin B6-dependent reaction, tetrahydrofolate accepts a one-carbon unit from serine, producing 5,10-methylenetetrahydrofolate and glycine.
5,10-Methylenetetrahydrofolate is required for the synthesis of nucleic acids. The figure shows the reduction of 5,10-methylenetetrahydrofolate to 5-methyltetrahydrofolate by a riboflavin (FAD)-dependent enzyme (methylenetetrahydrofolate reductase; MTHFR).
5-Methyltetrahydrofolate is required for the formation of methionine from homocysteine; this reaction also requires the cofactor vitamin B12 and betaine and releases dimethylglycine. Methionine, in the form of the methyl donor S-adenosylmethionine (SAM), is essential for many reactions, including DNA methylation.
The figure shows a methylation reaction catalyzed by a methyltransferase (shown as "X" converted to "methyl-X"), in which SAM donates a methyl group and is converted to S-adenosylhomocysteine (SAH). SAH is then converted to homocysteine, which enters the transsulfuration pathway and, through a vitamin B6-dependent reaction, is metabolized to cysteine.
Figure 3. Homocysteine metabolism
The figure illustrates a biochemical pathway of homocysteine metabolism. Homocysteine is methylated to form the essential amino acid methionine in two pathways. The reaction of homocysteine remethylation catalyzed by the vitamin B12-dependent methionine synthase captures a methyl group from the folate-dependent one-carbon pool (5-methyltetrahydrofolate), producing methionine and regenerating tetrahydrofolate. A second pathway requires betaine as a methyl donor for the methylation of homocysteine to form methionine; this reaction is catalyzed by betaine homocysteine methyltransferase, releasing dimethylglycine.
The figure also shows the catabolic pathway of homocysteine, known as the transsulfuration pathway. Here, homocysteine is converted to the amino acid cysteine via two vitamin B6 (PLP)-dependent enzymes: Cystathionine β-synthase catalyzes the condensation of homocysteine with serine to form cystathionine, and then converted by cystathionine γ-lyase into cysteine, α-ketobutyrate, and ammonia.
Referencias
- Food and Nutrition Board, Institute of Medicine. Folate. Dietary Reference Intakes: Thiamin, Riboflavin, Niacin, Vitamin B6, Folate, Vitamin B12, Pantothenic Acid, Biotin, and Choline. Washington, D.C.: National Academy Press; 1998:196-305. (National Academy Press)
- Choi SW, Mason JB. Folate and carcinogenesis: an integrated scheme. J Nutr. 2000;130(2):129-132. (PubMed)
- Bailey LB, Gregory JF, 3rd. Folate metabolism and requirements. J Nutr. 1999;129(4):779-782. (PubMed)
- Gerhard GT, Duell PB. Homocysteine and atherosclerosis. Curr Opin Lipidol. 1999;10(5):417-428. (PubMed)
- Jacques PF, Bostom AG, Wilson PW, Rich S, Rosenberg IH, Selhub J. Determinants of plasma total homocysteine concentration in the Framingham Offspring cohort. Am J Clin Nutr. 2001;73(3):613-621. (PubMed)
- Jacques PF, Kalmbach R, Bagley PJ, et al. The relationship between riboflavin and plasma total homocysteine in the Framingham Offspring cohort is influenced by folate status and the C677T transition in the methylenetetrahydrofolate reductase gene. J Nutr. 2002;132(2):283-288. (PubMed)
- McNulty H, Dowey le RC, Strain JJ, et al. Riboflavin lowers homocysteine in individuals homozygous for the MTHFR 677C->T polymorphism. Circulation. 2006;113(1):74-80. (PubMed)
- Verlinde PH, Oey I, Hendrickx ME, Van Loey AM, Temme EH. L-ascorbic acid improves the serum folate response to an oral dose of [6S]-5-methyltetrahydrofolic acid in healthy men. Eur J Clin Nutr. 2008;62(10):1224-1230. (PubMed)
- Lucock M, Yates Z, Boyd L, et al. Vitamin C-related nutrient-nutrient and nutrient-gene interactions that modify folate status. Eur J Nutr. 2013;52(2):569-582. (PubMed)
- Desmoulin SK, Hou Z, Gangjee A, Matherly LH. The human proton-coupled folate transporter: Biology and therapeutic applications to cancer. Cancer Biol Ther. 2012;13(14):1355-1373. (PubMed)
- Solanky N, Requena Jimenez A, D'Souza SW, Sibley CP, Glazier JD. Expression of folate transporters in human placenta and implications for homocysteine metabolism. Placenta. 2010;31(2):134-143. (PubMed)
- Halsted CH, Villanueva JA, Devlin AM, Chandler CJ. Metabolic interactions of alcohol and folate. J Nutr. 2002;132(8 Suppl):2367S-2372S. (PubMed)
- Pfeiffer CM, Sternberg MR, Schleicher RL, Rybak ME. Dietary supplement use and smoking are important correlates of biomarkers of water-soluble vitamin status after adjusting for sociodemographic and lifestyle variables in a representative sample of US adults. J Nutr. 2013;143(6):957S-965S. (PubMed)
- Stark KD, Pawlosky RJ, Sokol RJ, Hannigan JH, Salem N, Jr. Maternal smoking is associated with decreased 5-methyltetrahydrofolate in cord plasma. Am J Clin Nutr. 2007;85(3):796-802. (PubMed)
- Hutson JR, Stade B, Lehotay DC, Collier CP, Kapur BM. Folic acid transport to the human fetus is decreased in pregnancies with chronic alcohol exposure. PLoS One. 2012;7(5):e38057. (PubMed)
- Herbert V. Folic acid. In: Shils M, Olson JA, Shike M, Ross AC, eds. Modern Nutrition in Health and Disease. 9th ed. Baltimore: Lippincott Williams & Wilkins; 1999:433-446.
- Stabler SP. Clinical folate deficiency. In: Bailey LB, ed. Folate in Health and Disease. 2nd edition ed. Boca Raton, FL: CRC press, Taylor & Francis Group; 2010:409-428.
- Bailey LB. Dietary reference intakes for folate: the debut of dietary folate equivalents. Nutr Rev. 1998;56(10):294-299. (PubMed)
- Bailey LB, Gregory JF, 3rd. Polymorphisms of methylenetetrahydrofolate reductase and other enzymes: metabolic significance, risks and impact on folate requirement. J Nutr. 1999;129(5):919-922. (PubMed)
- Wilcken B, Bamforth F, Li Z, et al. Geographical and ethnic variation of the 677C>T allele of 5,10 methylenetetrahydrofolate reductase (MTHFR): findings from over 7000 newborns from 16 areas world wide. J Med Genet. 2003;40(8):619-625. (PubMed)
- Guenther BD, Sheppard CA, Tran P, Rozen R, Matthews RG, Ludwig ML. The structure and properties of methylenetetrahydrofolate reductase from Escherichia coli suggest how folate ameliorates human hyperhomocysteinemia. Nat Struct Biol. 1999;6(4):359-365. (PubMed)
- Molloy AM, Daly S, Mills JL, et al. Thermolabile variant of 5,10-methylenetetrahydrofolate reductase associated with low red-cell folates: implications for folate intake recommendations. Lancet. 1997;349(9065):1591-1593. (PubMed)
- Rozen R. Genetic predisposition to hyperhomocysteinemia: deficiency of methylenetetrahydrofolate reductase (MTHFR). Thromb Haemost. 1997;78(1):523-526. (PubMed)
- Kauwell GP, Wilsky CE, Cerda JJ, et al. Methylenetetrahydrofolate reductase mutation (677C-->T) negatively influences plasma homocysteine response to marginal folate intake in elderly women. Metabolism. 2000;49(11):1440-1443. (PubMed)
- Shane B. Folic acid, vitamin B-12, and vitamin B-6. In: Stipanuk M, ed. Biochemical and Physiological Aspects of Human Nutrition. Philadelphia: W.B. Saunders Co.; 2000:483-518.
- Eskes TK. Open or closed? A world of difference: a history of homocysteine research. Nutr Rev. 1998;56(8):236-244. (PubMed)
- Czeizel AE, Dudas I, Vereczkey A, Banhidy F. Folate deficiency and folic acid supplementation: the prevention of neural-tube defects and congenital heart defects. Nutrients. 2013;5(11):4760-4775. (PubMed)
- MRC Vitamin Study Research Group. Prevention of neural tube defects: results of the Medical Research Council Vitamin Study. Lancet. 1991;338(8760):131-137. (PubMed)
- Czeizel AE, Dudas I. Prevention of the first occurrence of neural-tube defects by periconceptional vitamin supplementation. N Engl J Med. 1992;327(26):1832-1835. (PubMed)
- Talaulikar VS, Arulkumaran S. Folic acid in obstetric practice: a review. Obstet Gynecol Surv. 2011;66(4):240-247. (PubMed)
- American College of Obstetricians and Gynecologists (ACOG). Neural tube defects. Washington, DC. 2003. Available at: http://www.guideline.gov/content.aspx?id=3994. Accessed 12/19/14.
- McNulty B, Pentieva K, Marshall B, et al. Women's compliance with current folic acid recommendations and achievement of optimal vitamin status for preventing neural tube defects. Hum Reprod. 2011;26(6):1530-1536. (PubMed)
- Nilsen RM, Vollset SE, Gjessing HK, et al. Patterns and predictors of folic acid supplement use among pregnant women: the Norwegian Mother and Child Cohort Study. Am J Clin Nutr. 2006;84(5):1134-1141. (PubMed)
- Ray JG, Singh G, Burrows RF. Evidence for suboptimal use of periconceptional folic acid supplements globally. BJOG. 2004;111(5):399-408. (PubMed)
- Quinlivan EP, Gregory JF, 3rd. Effect of food fortification on folic acid intake in the United States. Am J Clin Nutr. 2003;77(1):221-225. (PubMed)
- National Birth Defects Prevention Network. Neural Tube Defect Ascertainment Project. Available at: http://www.nbdpn.org/ntd_folic_acid_information.php. Accessed 12/16/14.
- Copp AJ, Stanier P, Greene ND. Neural tube defects: recent advances, unsolved questions, and controversies. Lancet Neurol. 2013;12(8):799-810. (PubMed)
- Yan L, Zhao L, Long Y, et al. Association of the maternal MTHFR C677T polymorphism with susceptibility to neural tube defects in offsprings: evidence from 25 case-control studies. PLoS One. 2012;7(10):e41689. (PubMed)
- De Marco P, Calevo MG, Moroni A, et al. Study of MTHFR and MS polymorphisms as risk factors for NTD in the Italian population. J Hum Genet. 2002;47(6):319-324. (PubMed)
- van der Put NM, Gabreels F, Stevens EM, et al. A second common mutation in the methylenetetrahydrofolate reductase gene: an additional risk factor for neural-tube defects? Am J Hum Genet. 1998;62(5):1044-1051. (PubMed)
- De Marco P, Calevo MG, Moroni A, et al. Reduced folate carrier polymorphism (80A-->G) and neural tube defects. Eur J Hum Genet. 2003;11(3):245-252. (PubMed)
- O'Leary VB, Mills JL, Parle-McDermott A, et al. Screening for new MTHFR polymorphisms and NTD risk. Am J Med Genet A. 2005;138A(2):99-106. (PubMed)
- Christensen B, Arbour L, Tran P, et al. Genetic polymorphisms in methylenetetrahydrofolate reductase and methionine synthase, folate levels in red blood cells, and risk of neural tube defects. Am J Med Genet. 1999;84(2):151-157. (PubMed)
- Relton CL, Wilding CS, Pearce MS, et al. Gene-gene interaction in folate-related genes and risk of neural tube defects in a UK population. J Med Genet. 2004;41(4):256-260. (PubMed)
- Brody LC, Conley M, Cox C, et al. A polymorphism, R653Q, in the trifunctional enzyme methylenetetrahydrofolate dehydrogenase/methenyltetrahydrofolate cyclohydrolase/formyltetrahydrofolate synthetase is a maternal genetic risk factor for neural tube defects: report of the Birth Defects Research Group. Am J Hum Genet. 2002;71(5):1207-1215. (PubMed)
- van der Put NM, van den Heuvel LP, Steegers-Theunissen RP, et al. Decreased methylene tetrahydrofolate reductase activity due to the 677C-->T mutation in families with spina bifida offspring. J Mol Med (Berl). 1996;74(11):691-694. (PubMed)
- Wilson A, Platt R, Wu Q, et al. A common variant in methionine synthase reductase combined with low cobalamin (vitamin B12) increases risk for spina bifida. Mol Genet Metab. 1999;67(4):317-323. (PubMed)
- Gilboa SM, Salemi JL, Nembhard WN, Fixler DE, Correa A. Mortality resulting from congenital heart disease among children and adults in the United States, 1999 to 2006. Circulation. 2010;122(22):2254-2263. (PubMed)
- van Beynum IM, Kapusta L, Bakker MK, den Heijer M, Blom HJ, de Walle HE. Protective effect of periconceptional folic acid supplements on the risk of congenital heart defects: a registry-based case-control study in the northern Netherlands. Eur Heart J. 2010;31(4):464-471. (PubMed)
- Yin M, Dong L, Zheng J, Zhang H, Liu J, Xu Z. Meta analysis of the association between MTHFR C677T polymorphism and the risk of congenital heart defects. Ann Hum Genet. 2012;76(1):9-16. (PubMed)
- Wang W, Wang Y, Gong F, Zhu W, Fu S. MTHFR C677T polymorphism and risk of congenital heart defects: evidence from 29 case-control and TDT studies. PLoS One. 2013;8(3):e58041. (PubMed)
- Badovinac RL, Werler MM, Williams PL, Kelsey KT, Hayes C. Folic acid-containing supplement consumption during pregnancy and risk for oral clefts: a meta-analysis. Birth Defects Res A Clin Mol Teratol. 2007;79(1):8-15. (PubMed)
- Wilcox AJ, Lie RT, Solvoll K, et al. Folic acid supplements and risk of facial clefts: national population based case-control study. BMJ. 2007;334(7591):464. (PubMed)
- Boyles AL, Wilcox AJ, Taylor JA, et al. Folate and one-carbon metabolism gene polymorphisms and their associations with oral facial clefts. Am J Med Genet A. 2008;146A(4):440-449. (PubMed)
- Boyles AL, Wilcox AJ, Taylor JA, et al. Oral facial clefts and gene polymorphisms in metabolism of folate/one-carbon and vitamin A: a pathway-wide association study. Genet Epidemiol. 2009;33(3):247-255. (PubMed)
- Luo YL, Cheng YL, Ye P, Wang W, Gao XH, Chen Q. Association between MTHFR polymorphisms and orofacial clefts risk: a meta-analysis. Birth Defects Res A Clin Mol Teratol. 2012;94(4):237-244. (PubMed)
- Wilcox AJ. On the importance--and the unimportance--of birthweight. Int J Epidemiol. 2001;30(6):1233-1241. (PubMed)
- Fekete K, Berti C, Trovato M, et al. Effect of folate intake on health outcomes in pregnancy: a systematic review and meta-analysis on birth weight, placental weight and length of gestation. Nutr J. 2012;11:75. (PubMed)
- Baker PN, Wheeler SJ, Sanders TA, et al. A prospective study of micronutrient status in adolescent pregnancy. Am J Clin Nutr. 2009;89(4):1114-1124. (PubMed)
- Lee HA, Park EA, Cho SJ, et al. Mendelian randomization analysis of the effect of maternal homocysteine during pregnancy, as represented by maternal MTHFR C677T genotype, on birth weight. J Epidemiol. 2013;23(5):371-375. (PubMed)
- Scholl TO, Johnson WG. Folic acid: influence on the outcome of pregnancy. Am J Clin Nutr. 2000;71(5 Suppl):1295S-1303S. (PubMed)
- Vollset SE, Refsum H, Irgens LM, et al. Plasma total homocysteine, pregnancy complications, and adverse pregnancy outcomes: the Hordaland Homocysteine study. Am J Clin Nutr. 2000;71(4):962-968. (PubMed)
- Wang XM, Wu HY, Qiu XJ. Methylenetetrahydrofolate reductase (MTHFR) gene C677T polymorphism and risk of preeclampsia: an updated meta-analysis based on 51 studies. Arch Med Res. 2013;44(3):159-168. (PubMed)
- Wen SW, Champagne J, Rennicks White R, et al. Effect of folic acid supplementation in pregnancy on preeclampsia: the folic acid clinical trial study. J Pregnancy. 2013;2013:294312. (PubMed)
- Lassi ZS, Salam RA, Haider BA, Bhutta ZA. Folic acid supplementation during pregnancy for maternal health and pregnancy outcomes. Cochrane Database Syst Rev. 2013;3:CD006896. (PubMed)
- Schmidt RJ, Tancredi DJ, Ozonoff S, et al. Maternal periconceptional folic acid intake and risk of autism spectrum disorders and developmental delay in the CHARGE (CHildhood Autism Risks from Genetics and Environment) case-control study. Am J Clin Nutr. 2012;96(1):80-89. (PubMed)
- Crider KS, Cordero AM, Qi YP, Mulinare J, Dowling NF, Berry RJ. Prenatal folic acid and risk of asthma in children: a systematic review and meta-analysis. Am J Clin Nutr. 2013;98(5):1272-1281. (PubMed)
- Brown SB, Reeves KW, Bertone-Johnson ER. Maternal folate exposure in pregnancy and childhood asthma and allergy: a systematic review. Nutr Rev. 2014;72(1):55-64. (PubMed)
- Ding R, Lin S, Chen D. The association of cystathionine β synthase (CBS) T833C polymorphism and the risk of stroke: a meta-analysis. J Neurol Sci. 2012;312(1-2):26-30. (PubMed)
- Seshadri N, Robinson K. Homocysteine, B vitamins, and coronary artery disease. Med Clin North Am. 2000;84(1):215-237. (PubMed)
- Wald DS, Law M, Morris JK. Homocysteine and cardiovascular disease: evidence on causality from a meta-analysis. BMJ. 2002;325(7374):1202. (PubMed)
- Homocysteine Studies Collaboration. Homocysteine and risk of ischemic heart disease and stroke: a meta-analysis. JAMA. 2002;288(16):2015-2022. (PubMed)
- Clarke R, Halsey J, Bennett D, Lewington S. Homocysteine and vascular disease: review of published results of the homocysteine-lowering trials. J Inherit Metab Dis. 2011;34(1):83-91. (PubMed)
- Huang T, Chen Y, Yang B, Yang J, Wahlqvist ML, Li D. Meta-analysis of B vitamin supplementation on plasma homocysteine, cardiovascular and all-cause mortality. Clin Nutr. 2012;31(4):448-454. (PubMed)
- Voutilainen S, Rissanen TH, Virtanen J, Lakka TA, Salonen JT. Low dietary folate intake is associated with an excess incidence of acute coronary events: The Kuopio Ischemic Heart Disease Risk Factor Study. Circulation. 2001;103(22):2674-2680. (PubMed)
- Brattstrom L. Vitamins as homocysteine-lowering agents. J Nutr. 1996;126(4 Suppl):1276S-1280S. (PubMed)
- Rader JI. Folic acid fortification, folate status and plasma homocysteine. J Nutr. 2002;132(8 Suppl):2466S-2470S. (PubMed)
- Homocysteine Lowering Trialists' Collaboration. Dose-dependent effects of folic acid on blood concentrations of homocysteine: a meta-analysis of the randomized trials. Am J Clin Nutr. 2005;82(4):806-812. (PubMed)
- Malinow MR, Bostom AG, Krauss RM. Homocyst(e)ine, diet, and cardiovascular diseases: a statement for healthcare professionals from the Nutrition Committee, American Heart Association. Circulation. 1999;99(1):178-182. (PubMed)
- van Meurs JB, Pare G, Schwartz SM, et al. Common genetic loci influencing plasma homocysteine concentrations and their effect on risk of coronary artery disease. Am J Clin Nutr. 2013;98(3):668-676. (PubMed)
- Holmes MV, Newcombe P, Hubacek JA, et al. Effect modification by population dietary folate on the association between MTHFR genotype, homocysteine, and stroke risk: a meta-analysis of genetic studies and randomised trials. Lancet. 2011;378(9791):584-594. (PubMed)
- Clarke R, Bennett DA, Parish S, et al. Homocysteine and coronary heart disease: meta-analysis of MTHFR case-control studies, avoiding publication bias. PLoS Med. 2012;9(2):e1001177. (PubMed)
- Ji Y, Tan S, Xu Y, et al. Vitamin B supplementation, homocysteine levels, and the risk of cerebrovascular disease: A meta-analysis. Neurology. 2013;81(15):1298-1307. (PubMed)
- Zhang C, Chi FL, Xie TH, Zhou YH. Effect of B-vitamin supplementation on stroke: a meta-analysis of randomized controlled trials. PLoS One. 2013;8(11):e81577. (PubMed)
- Mosca L, Benjamin EJ, Berra K, et al. Effectiveness-based guidelines for the prevention of cardiovascular disease in women--2011 update: a guideline from the American Heart Association. J Am Coll Cardiol. 2011;57(12):1404-1423. (PubMed)
- Wang X, Qin X, Demirtas H, et al. Efficacy of folic acid supplementation in stroke prevention: a meta-analysis. Lancet. 2007;369(9576):1876-1882. (PubMed)
- Yang Q, Botto LD, Erickson JD, et al. Improvement in stroke mortality in Canada and the United States, 1990 to 2002. Circulation. 2006;113(10):1335-1343. (PubMed)
- Lorenz MW, Markus HS, Bots ML, Rosvall M, Sitzer M. Prediction of clinical cardiovascular events with carotid intima-media thickness: a systematic review and meta-analysis. Circulation. 2007;115(4):459-467. (PubMed)
- Qin X, Xu M, Zhang Y, et al. Effect of folic acid supplementation on the progression of carotid intima-media thickness: a meta-analysis of randomized controlled trials. Atherosclerosis. 2012;222(2):307-313. (PubMed)
- de Bree A, van Mierlo LA, Draijer R. Folic acid improves vascular reactivity in humans: a meta-analysis of randomized controlled trials. Am J Clin Nutr. 2007;86(3):610-617. (PubMed)
- McNeil CJ, Beattie JH, Gordon MJ, Pirie LP, Duthie SJ. Nutritional B vitamin deficiency disrupts lipid metabolism causing accumulation of proatherogenic lipoproteins in the aorta adventitia of ApoE null mice. Mol Nutr Food Res. 2012;56(7):1122-1130. (PubMed)
- Blount BC, Mack MM, Wehr CM, et al. Folate deficiency causes uracil misincorporation into human DNA and chromosome breakage: implications for cancer and neuronal damage. Proc Natl Acad Sci U S A. 1997;94(7):3290-3295. (PubMed)
- Narayanan S, McConnell J, Little J, et al. Associations between two common variants C677T and A1298C in the methylenetetrahydrofolate reductase gene and measures of folate metabolism and DNA stability (strand breaks, misincorporated uracil, and DNA methylation status) in human lymphocytes in vivo. Cancer Epidemiol Biomarkers Prev. 2004;13(9):1436-1443. (PubMed)
- Rahman L, Voeller D, Rahman M, et al. Thymidylate synthase as an oncogene: a novel role for an essential DNA synthesis enzyme. Cancer Cell. 2004;5(4):341-351. (PubMed)
- Hubner RA, Liu JF, Sellick GS, Logan RF, Houlston RS, Muir KR. Thymidylate synthase polymorphisms, folate and B-vitamin intake, and risk of colorectal adenoma. Br J Cancer. 2007;97(10):1449-1456. (PubMed)
- Desmoulin SK, Wang L, Polin L, et al. Functional loss of the reduced folate carrier enhances the antitumor activities of novel antifolates with selective uptake by the proton-coupled folate transporter. Mol Pharmacol. 2012;82(4):591-600. (PubMed)
- Crider KS, Yang TP, Berry RJ, Bailey LB. Folate and DNA methylation: a review of molecular mechanisms and the evidence for folate's role. Adv Nutr. 2012;3(1):21-38. (PubMed)
- Butrum RR, Clifford CK, Lanza E. NCI dietary guidelines: rationale. Am J Clin Nutr. 1988;48(3 Suppl):888-895. (PubMed)
- Crider KS, Bailey LB, Berry RJ. Folic acid food fortification-its history, effect, concerns, and future directions. Nutrients. 2011;3(3):370-384. (PubMed)
- Qin X, Cui Y, Shen L, et al. Folic acid supplementation and cancer risk: A meta-analysis of randomized controlled trials. Int J Cancer. 2013;133(5):1033-1041. (PubMed)
- Vollset SE, Clarke R, Lewington S, et al. Effects of folic acid supplementation on overall and site-specific cancer incidence during the randomised trials: meta-analyses of data on 50,000 individuals. Lancet. 2013;381(9871):1029-1036. (PubMed)
- Kim DH, Smith-Warner SA, Spiegelman D, et al. Pooled analyses of 13 prospective cohort studies on folate intake and colon cancer. Cancer Causes Control. 2010;21(11):1919-1930. (PubMed)
- Gibson TM, Weinstein SJ, Pfeiffer RM, et al. Pre- and postfortification intake of folate and risk of colorectal cancer in a large prospective cohort study in the United States. Am J Clin Nutr. 2011;94(4):1053-1062. (PubMed)
- Stevens VL, McCullough ML, Sun J, Jacobs EJ, Campbell PT, Gapstur SM. High levels of folate from supplements and fortification are not associated with increased risk of colorectal cancer. Gastroenterology. 2011;141(1):98-105, 105 e101. (PubMed)
- Zschabitz S, Cheng TY, Neuhouser ML, et al. B vitamin intakes and incidence of colorectal cancer: results from the Women's Health Initiative Observational Study cohort. Am J Clin Nutr. 2013;97(2):332-343. (PubMed)
- Keum N, Giovannucci EL. Folic acid fortification and colorectal cancer risk. Am J Prev Med. 2014;46(3 Suppl 1):S65-72. (PubMed)
- Kennedy DA, Stern SJ, Moretti M, et al. Folate intake and the risk of colorectal cancer: a systematic review and meta-analysis. Cancer Epidemiol. 2011;35(1):2-10. (PubMed)
- Kim YI. Folate: a magic bullet or a double edged sword for colorectal cancer prevention? Gut. 2006;55(10):1387-1389. (PubMed)
- Paspatis GA, Kalafatis E, Oros L, Xourgias V, Koutsioumpa P, Karamanolis DG. Folate status and adenomatous colonic polyps. A colonoscopically controlled study. Dis Colon Rectum. 1995;38(1):64-67; discussion 67-68. (PubMed)
- Jaszewski R, Misra S, Tobi M, et al. Folic acid supplementation inhibits recurrence of colorectal adenomas: a randomized chemoprevention trial. World J Gastroenterol. 2008;14(28):4492-4498. (PubMed)
- Wu K, Platz EA, Willett WC, et al. A randomized trial on folic acid supplementation and risk of recurrent colorectal adenoma. Am J Clin Nutr. 2009;90(6):1623-1631. (PubMed)
- Logan RF, Grainge MJ, Shepherd VC, Armitage NC, Muir KR. Aspirin and folic acid for the prevention of recurrent colorectal adenomas. Gastroenterology. 2008;134(1):29-38. (PubMed)
- Figueiredo JC, Mott LA, Giovannucci E, et al. Folic acid and prevention of colorectal adenomas: a combined analysis of randomized clinical trials. Int J Cancer. 2011;129(1):192-203. (PubMed)
- Kennedy DA, Stern SJ, Matok I, et al. Folate intake, MTHFR polymorphisms, and the risk of colorectal cancer: a systematic review and meta-analysis. J Cancer Epidemiol. 2012;2012:952508. (PubMed)
- Ding W, Zhou DL, Jiang X, Lu LS. Methionine synthase A2756G polymorphism and risk of colorectal adenoma and cancer: evidence based on 27 studies. PLoS One. 2013;8(4):e60508. (PubMed)
- Nan H, Lee JE, Rimm EB, Fuchs CS, Giovannucci EL, Cho E. Prospective study of alcohol consumption and the risk of colorectal cancer before and after folic acid fortification in the United States. Ann Epidemiol. 2013;23(9):558-563. (PubMed)
- Slattery ML, Potter JD, Samowitz W, Schaffer D, Leppert M. Methylenetetrahydrofolate reductase, diet, and risk of colon cancer. Cancer Epidemiol Biomarkers Prev. 1999;8(6):513-518. (PubMed)
- Ma J, Stampfer MJ, Giovannucci E, et al. Methylenetetrahydrofolate reductase polymorphism, dietary interactions, and risk of colorectal cancer. Cancer Res. 1997;57(6):1098-1102. (PubMed)
- Larsson SC, Giovannucci E, Wolk A. Folate and risk of breast cancer: a meta-analysis. J Natl Cancer Inst. 2007;99(1):64-76. (PubMed)
- Liu M, Cui LH, Ma AG, Li N, Piao JM. Lack of effects of dietary folate intake on risk of breast cancer: an updated meta-analysis of prospective studies. Asian Pac J Cancer Prev. 2014;15(5):2323-2328. (PubMed)
- Brooks PJ, Zakhari S. Moderate alcohol consumption and breast cancer in women: from epidemiology to mechanisms and interventions. Alcohol Clin Exp Res. 2013;37(1):23-30. (PubMed)
- Rohan TE, Jain MG, Howe GR, Miller AB. Dietary folate consumption and breast cancer risk. J Natl Cancer Inst. 2000;92(3):266-269. (PubMed)
- Sellers TA, Kushi LH, Cerhan JR, et al. Dietary folate intake, alcohol, and risk of breast cancer in a prospective study of postmenopausal women. Epidemiology. 2001;12(4):420-428. (PubMed)
- Zhang S, Hunter DJ, Hankinson SE, et al. A prospective study of folate intake and the risk of breast cancer. JAMA. 1999;281(17):1632-1637. (PubMed)
- Tjonneland A, Christensen J, Olsen A, et al. Alcohol intake and breast cancer risk: the European Prospective Investigation into Cancer and Nutrition (EPIC). Cancer Causes Control. 2007;18(4):361-373. (PubMed)
- Bassett JK, Baglietto L, Hodge AM, et al. Dietary intake of B vitamins and methionine and breast cancer risk. Cancer Causes Control. 2013;24(8):1555-1563. (PubMed)
- Wang J, Wang B, Bi J, Di J. The association between two polymorphisms in the TYMS gene and breast cancer risk: a meta-analysis. Breast Cancer Res Treat. 2011;128(1):203-209. (PubMed)
- Weiner AS, Boyarskikh UA, Voronina EN, et al. Polymorphisms in the folate-metabolizing genes MTR, MTRR, and CBS and breast cancer risk. Cancer Epidemiol. 2012;36(2):e95-e100. (PubMed)
- Linabery AM, Johnson KJ, Ross JA. Childhood cancer incidence trends in association with US folic acid fortification (1986-2008). Pediatrics. 2012;129(6):1125-1133. (PubMed)
- Milne E, Royle JA, Miller M, et al. Maternal folate and other vitamin supplementation during pregnancy and risk of acute lymphoblastic leukemia in the offspring. Int J Cancer. 2010;126(11):2690-2699. (PubMed)
- Yan J, Yin M, Dreyer ZE, et al. A meta-analysis of MTHFR C677T and A1298C polymorphisms and risk of acute lymphoblastic leukemia in children. Pediatr Blood Cancer. 2012;58(4):513-518. (PubMed)
- Alzheimer's Association. 2013 Alzheimer's Disease Fact and Figures. Alzheimer's & Dementia. 9(2). Available at: http://www.alz.org/downloads/facts_figures_2013.pdf. Accessed 9/9/13.
- Hughes TF, Andel R, Small BJ, et al. Midlife fruit and vegetable consumption and risk of dementia in later life in Swedish twins. Am J Geriatr Psychiatry. 2010;18(5):413-420. (PubMed)
- Weir DG, Scott JM. Brain function in the elderly: role of vitamin B12 and folate. Br Med Bull. 1999;55(3):669-682. (PubMed)
- Faux NG, Ellis KA, Porter L, et al. Homocysteine, vitamin B12, and folic acid levels in Alzheimer's disease, mild cognitive impairment, and healthy elderly: baseline characteristics in subjects of the Australian Imaging Biomarker Lifestyle study. J Alzheimers Dis. 2011;27(4):909-922. (PubMed)
- Van Dam F, Van Gool WA. Hyperhomocysteinemia and Alzheimer's disease: A systematic review. Arch Gerontol Geriatr. 2009;48(3):425-430. (PubMed)
- Morris MC, Evans DA, Bienias JL, et al. Dietary folate and vitamin B12 intake and cognitive decline among community-dwelling older persons. Arch Neurol. 2005;62(4):641-645. (PubMed)
- Morris MC, Evans DA, Schneider JA, Tangney CC, Bienias JL, Aggarwal NT. Dietary folate and vitamins B-12 and B-6 not associated with incident Alzheimer's disease. J Alzheimers Dis. 2006;9(4):435-443. (PubMed)
- Wald DS, Kasturiratne A, Simmonds M. Serum homocysteine and dementia: meta-analysis of eight cohort studies including 8669 participants. Alzheimers Dement. 2011;7(4):412-417. (PubMed)
- Ho RC, Cheung MW, Fu E, et al. Is high homocysteine level a risk factor for cognitive decline in elderly? A systematic review, meta-analysis, and meta-regression. Am J Geriatr Psychiatry. 2011;19(7):607-617. (PubMed)
- Nilforooshan R, Broadbent D, Weaving G, et al. Homocysteine in Alzheimer's disease: role of dietary folate, vitamin B6 and B12. Int J Geriatr Psychiatry. 2011;26(8):876-877. (PubMed)
- Wald DS, Kasturiratne A, Simmonds M. Effect of folic acid, with or without other B vitamins, on cognitive decline: meta-analysis of randomized trials. Am J Med. 2010;123(6):522-527 e522. (PubMed)
- Ford AH, Almeida OP. Effect of homocysteine lowering treatment on cognitive function: a systematic review and meta-analysis of randomized controlled trials. J Alzheimers Dis. 2012;29(1):133-149. (PubMed)
- Nachum-Biala Y, Troen AM. B-vitamins for neuroprotection: narrowing the evidence gap. Biofactors. 2012;38(2):145-150. (PubMed)
- Smith AD, Smith SM, de Jager CA, et al. Homocysteine-lowering by B vitamins slows the rate of accelerated brain atrophy in mild cognitive impairment: a randomized controlled trial. PLoS One. 2010;5(9):e12244. (PubMed)
- Douaud G, Refsum H, de Jager CA, et al. Preventing Alzheimer's disease-related gray matter atrophy by B-vitamin treatment. Proc Natl Acad Sci U S A. 2013;110(23):9523-9528. (PubMed)
- Watkins D, Rosenblatt DS. Update and new concepts in vitamin responsive disorders of folate transport and metabolism. J Inherit Metab Dis. 2012;35(4):665-670. (PubMed)
- Zhao R, Min SH, Qiu A, et al. The spectrum of mutations in the PCFT gene, coding for an intestinal folate transporter, that are the basis for hereditary folate malabsorption. Blood. 2007;110(4):1147-1152. (PubMed)
- Borzutzky A, Crompton B, Bergmann AK, et al. Reversible severe combined immunodeficiency phenotype secondary to a mutation of the proton-coupled folate transporter. Clin Immunol. 2009;133(3):287-294. (PubMed)
- Sofer Y, Harel L, Sharkia M, Amir J, Schoenfeld T, Straussberg R. Neurological manifestations of folate transport defect: case report and review of the literature. J Child Neurol. 2007;22(6):783-786. (PubMed)
- Diop-Bove N, Kronn D, Goldman ID. Hereditary folate malabsorption. In: Pagon RA, Adam MP, Bird TD, Dolan CR, Fong CT, Stephens K, eds. GeneReviews™ [Internet]. Seattle, WA: University of Washington, Seattle; 2008. (PubMed)
- Frye RE, Sequeira JM, Quadros EV, James SJ, Rossignol DA. Cerebral folate receptor autoantibodies in autism spectrum disorder. Mol Psychiatry. 2013;18(3):369-381. (PubMed)
- Grapp M, Just IA, Linnankivi T, et al. Molecular characterization of folate receptor 1 mutations delineates cerebral folate transport deficiency. Brain. 2012;135(Pt 7):2022-2031. (PubMed)
- Ramaekers VT, Blau N, Sequeira JM, Nassogne MC, Quadros EV. Folate receptor autoimmunity and cerebral folate deficiency in low-functioning autism with neurological deficits. Neuropediatrics. 2007;38(6):276-281. (PubMed)
- Ramaekers VT, Hausler M, Opladen T, Heimann G, Blau N. Psychomotor retardation, spastic paraplegia, cerebellar ataxia and dyskinesia associated with low 5-methyltetrahydrofolate in cerebrospinal fluid: a novel neurometabolic condition responding to folinic acid substitution. Neuropediatrics. 2002;33(6):301-308. (PubMed)
- Banka S, Blom HJ, Walter J, et al. Identification and characterization of an inborn error of metabolism caused by dihydrofolate reductase deficiency. Am J Hum Genet. 2011;88(2):216-225. (PubMed)
- Cario H, Smith DE, Blom H, et al. Dihydrofolate reductase deficiency due to a homozygous DHFR mutation causes megaloblastic anemia and cerebral folate deficiency leading to severe neurologic disease. Am J Hum Genet. 2011;88(2):226-231. (PubMed)
- Pfeiffer CM, Hughes JP, Lacher DA, et al. Estimation of trends in serum and RBC folate in the US population from pre- to postfortification using assay-adjusted data from the NHANES 1988-2010. J Nutr. 2012;142(5):886-893. (PubMed)
- Folate. In: Hendler SS, Rorvik, D.R., ed. PDR for Nutritional Supplements. 2nd ed. Montvale: Physicians' Desk Reference Inc.; 2008.
- Wiesinger H, Eydeler U, Richard F, et al. Bioequivalence evaluation of a folate-supplemented oral contraceptive containing ethinylestradiol/drospirenone/levomefolate calcium versus ethinylestradiol/drospirenone and levomefolate calcium alone. Clin Drug Investig. 2012;32(10):673-684. (PubMed)
- Tinker SC, Cogswell ME, Devine O, Berry RJ. Folic acid intake among US women aged 15-44 years, National Health and Nutrition Examination Survey, 2003-2006. Am J Prev Med. 2010;38(5):534-542. (PubMed)
- Kelly P, McPartlin J, Goggins M, Weir DG, Scott JM. Unmetabolized folic acid in serum: acute studies in subjects consuming fortified food and supplements. Am J Clin Nutr. 1997;65(6):1790-1795. (PubMed)
- Morris MS, Jacques PF, Rosenberg IH, Selhub J. Folate and vitamin B-12 status in relation to anemia, macrocytosis, and cognitive impairment in older Americans in the age of folic acid fortification. Am J Clin Nutr. 2007;85(1):193-200. (PubMed)
- Morris MS, Jacques PF, Rosenberg IH, Selhub J. Circulating unmetabolized folic acid and 5-methyltetrahydrofolate in relation to anemia, macrocytosis, and cognitive test performance in American seniors. Am J Clin Nutr. 2010;91(6):1733-1744. (PubMed)
- Troen AM, Mitchell B, Sorensen B, et al. Unmetabolized folic acid in plasma is associated with reduced natural killer cell cytotoxicity among postmenopausal women. J Nutr. 2006;136(1):189-194. (PubMed)
- Tam C, O'Connor D, Koren G. Circulating unmetabolized folic acid: relationship to folate status and effect of supplementation. Obstet Gynecol Int. 2012;2012:485179. (PubMed)
- Apeland T, Mansoor MA, Strandjord RE. Antiepileptic drugs as independent predictors of plasma total homocysteine levels. Epilepsy Res. 2001;47(1-2):27-35. (PubMed)
- Wilson SM, Bivins BN, Russell KA, Bailey LB. Oral contraceptive use: impact on folate, vitamin B(6), and vitamin B(1)(2) status. Nutr Rev. 2011;69(10):572-583. (PubMed)
Exclusión de Responsabilidades
El Centro de Informaciones sobre Micronutrientes del Instituto Linus Pauling entrega información científica relacionado con aspectos de importancia en salud pública de factores dietéticos, suplementos, comestibles y bebidas para el público en general. Esta información se entrega con el entendimiento que ni el autor ni la imprenta están entregando consejos de naturaleza médica, psicológica o nutricional. La información no debe ser usada para reemplazar la consulta con profesionales competentes en el área de cuidado de salud o de nutrición.
La información entregada con relación a factores y suplementos dietéticos, comestibles, y bebestibles, contenida en este sitio web, no cubre todos los usos, acciones, precauciones, efectos secundarios, e interacciones posibles. No debe ser considerado como consejo nutricional o médico para resolver problemas individuales. La responsabilidad legal por las acciones individuales u omisiones que se derivan del uso de este sitio está expresamente excluida.
No se puede copiar, modificar, distribuir, transmitir, realizar, publicar o vender ninguno de los materiales sujetos a derechos de autoría de este sitio web. Puede enlazar a este sitio web, pero debe incluir la siguiente declaración:
"Este enlace lleva a un sitio web proporcionado por el Instituto Linus Pauling en la Universidad Estatal de Oregon. [Su nombre] no está afiliado ni respaldado por el Instituto Linus Pauling o la Universidad Estatal de Oregon."