要約
-
ビタミンB5としても知られるパントテン酸は水溶性ビタミンで、補酵素Aの合成における前駆体である。補酵素Aは、生命を支える多くの生化学的反応に必須である。また、補酵素Aのホスホパンテテイニル基部分は、脂肪酸の合成に関わるアシルキャリアタンパク質を含むいくつかのタンパク質の生物学的活性に必要である。(詳細はこちら)
- パントテン酸は、すべての形態の生物に必須である。それは植物および動物由来の食物にあまねく含まれ、食事による欠乏症は非常に稀である。(詳細はこちら)
-
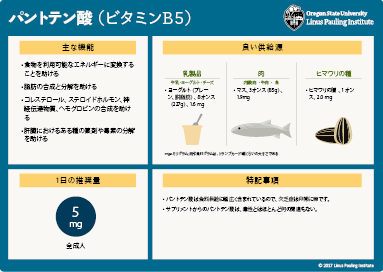
米国医学研究所の食品栄養委員会は、パントテン酸の推定1日平均摂取量に基づいて、5mg/日という成人の目安量(AI)を設定した。 (詳細はこちら)
- 限定的な介入研究によるエビデンスから、パントテン酸および/またはパントテノール(アルコール類似体)が皮膚の創傷治癒過程を改善させるかもしれないことが示唆される。今なお追加的なより大規模の研究が認められている。 (詳細はこちら)
- パントテン酸の誘導体であるパンテチンによる高用量の治療で、血清コレステロールや脂質濃度が下がることが示されてきた。パンテチンによる治療は耐用性が高いようであるが、医学的監督が欠かせない。 (詳細はこちら)
- パントテン酸の豊富な食品は、動物の臓器(肝臓や腎臓)、魚、貝、牛乳製品、卵、アボカド、豆類、マッシュルーム、およびサツマイモなどである。 (詳細はこちら)
- 食事性およびサプリメントからのパントテン酸は毒性とほとんど何の関連もないので、許容上限摂取量(UL)が設定されていないほどである。 (詳細はこちら)
ビタミンB5としても知られるパントテン酸は、すべての形態の生命に必須である(1)。パントテン酸は、多数の化学反応に非常に重要な補酵素である補酵素Aの形態で生物の全部門に見られる(2)。
機能
パントテン酸補助因子の合成
補酵素A
パントテン酸は、生命を支える多様な生化学反応に必須の補酵素である補酵素A(CoA、図1参照)の生合成における前駆体である(下記参照)。パントテン酸キナーゼII(PanK2)は、パントテン酸から4'-ホスホパントテン酸へのリン酸化の初期段階を触媒作用する。補酵素Aとその誘導体は4'-ホスホパントテン酸の合成を阻害するが、脂肪酸のミトコンドリアへの輸送に必要なカルニチンによってこの阻害は保留可能である(3)。この生合成経路におけるその後の反応には、中間体である4'-ホスホパンテテインの合成や、補酵素Aから4'-ホスホパンテテインへの再利用などがある(図1参照)。

4'-ホスホパンテテイン
補酵素Aの4'-ホスホパンテテイニル基部分は、生物学的活性のために4'-ホスホパンテテインが必須の補助因子である酵素類へと移送可能である(4'-ホスホパンテテイニル化の項参照)。
補助因子および補助基質機能
補酵素A
補酵素Aはアシル基と反応し、アセチルCoA、スクシニルCoA、マロニルCoA、および3-ヒドロキシ-3-メチルグルタリル(HMG)-CoAなどのチオエステル誘導体を生じる。補酵素Aとそのアシル誘導体は、食事性の脂肪、炭水化物、およびタンパク質の分解によってエネルギーを作り出す反応に必要である。また、アセチルCoAとスクシニルCoAの形態の補酵素Aはクエン酸回路や、必須脂肪、コレステロール、ステロイドホルモン、ビタミンAとD、および神経伝達物質のアセチルコリンの合成、さらに脂肪酸β酸化経路に関わっている。補酵素Aの誘導体はまた、メラトニンというホルモンの合成、およびヘムと呼ばれるヘモグロビン成分に必要である。加えて、肝臓での多数の薬剤や毒素の代謝に補酵素Aが必要である(4)。
補酵素Aは、そのアセチル化反応における役割から命名された。体内のアセチル化されたタンパク質の大部分は、補酵素Aのチオエステル誘導体であるアセチルCoAによって提供されたアセテート基を加えることによって修飾される。タンパク質のアセチル化はタンパク質の全体的な電荷を変化させ、その三次元的構造を変え、その機能まで変える可能性がある。たとえば、アセチル化は脳下垂体が生成するものも含めたペプチドホルモンの活性を調整するメカニズムである(5)。またタンパク質のアセチル化は、その他の翻訳後修飾と同様に、多くのシグナル(信号)分子、転写因子、および酵素の細胞内局在化、機能、半減期を調整することが示されている。特にヒストンのアセチル化は転写(すなわちmRNAの合成)を促進することで遺伝子発現の調整に役割を果たすが、脱アセチル化されたヒストンは通常、クロマチン凝縮や遺伝子発現抑制に関連がある。ヒストンのアセチル化はクロマチンの構造変化につながることがわかっており、これはDNAとタンパク質、およびタンパク質とタンパク質同士の相互作用に影響する。ヒストンのアセチル化標識とその他の転写後修飾のクロストーク(相互干渉)はまた、その後転写される遺伝子のプロモーターへの転写調整因子の補充も促す((6)の文献でレビュー)。
最後に、多くのシグナル分子は補酵素Aによって提供された長鎖脂肪酸の付加によって修飾される。これらの修飾はタンパク質のアシル化として知られ、細胞シグナル伝達経路に中心的役割を果たす(4)。
4'-ホスホパンテテイニル化
特定の多酵素複合体は、秩序だったやり方でいくつかの反応を実施する必要があるが、4'-ホスホパンテテインアームの「キャリア」領域(またはタンパク質)への共有結合を必要とするかもしれない。このキャリア領域は、様々な酵素反応の進行中に基質または反応中間体を保持する。哺乳類では、補酵素Aから特定のキャリア領域の保存セリン残基への4'-ホスホパンテテイニル基部分の輸送は、唯一ホスホパンテテイニル転移酵素によって触媒作用がなされる(7)。4'-ホスホパンテテイニル化は、アポ酵素を十分に活性のあるホロ酵素に変換するために必要である(下記参照)。
アシルキャリアタンパク質
脂質は正常な生理学的機能に必須の脂肪分子で、あまたある中にスフィンゴ脂質(神経伝達を強化するミエリン鞘の必須成分)、リン脂質(細胞膜の重要な構造成分)、および脂肪酸を含む。脂肪酸合成酵素(FAS)は、脂肪酸の合成を触媒する多酵素複合体である。FAS複合体の中でも、アシルキャリアタンパク質(ACP)はキャリアタンパク質としての活性のために4'-ホスホパンテテインの形態でパントテン酸を必要とする(3)。ACPにとっての4'-ホスホパンテテイニル基部分のような基は、補欠分子族と呼ばれる。補欠分子族はアミノ酸を含まず、いくつかのタンパク質の生物学的活性に必要な強固に結合した補助因子である(図2参照)。アセチルCoA、マロニルCoA、およびACPはどれも、サイトゾルにおける脂肪酸の合成に必要である。脂肪酸の合成中に、アセチルCoAとマロニルCoAのアシル基は、ACPの4'-ホスホパンテテイニル基部分のスルフヒドリル基(-SH)に輸送される。補欠分子族は、増殖中の脂肪酸鎖をI型FAS複合体の酵素的中心部のそれぞれに輸送するためのフレキシブル(可撓性)アームとして使用される。ミトコンドリアでは、4'-ホスホパンテテインはミトコンドリアのII型FAS複合体にあるACP相同体に対する補欠分子族としても作用する(8)。

10-ホルミルテトラヒドロ葉酸脱水素酵素
10-ホルミルテトラヒドロ葉酸脱水素酵素(FDH)は、10-ホルミルテトラヒドロ葉酸が核酸およびアミノ酸の代謝に不可欠の補助因子であるテトラヒドロ葉酸へと変換する際に触媒作用をする(図3参照)。ACPと同様に、FDHは自身の生物学的活性のために4'-ホスホパンテテイン補欠分子族を必要とする。補欠分子族は、FDHの2つの触媒領域の活性を結びつけるスイング(揺動)アームとして働く(9,10)。ミトコンドリアのFDHの相同体も、生物学的に活性を持つために4'-ホスホパンテテイニル化を必要とする(11)。

アミノアジピン酸セミアルデヒド合成酵素
4'-ホスホパンテテイニル化は、アポ酵素であるα-アミノアジピン酸セミアルデヒド合成酵素(AASS)の生物学的活性に必要である。AASSは、ヒトにとっての必須アミノ酸であるリジンをミトコンドリアで分解する経路における初期反応を触媒する。AASSは2つの触媒領域でできている。リジンケトグルタル酸還元酵素領域がまずリジンからサッカロピンへの変換を触媒する。サッカロピンはさらに、サッカロピン脱水素酵素領域によって触媒される反応においてα-アミノアジピン酸セミアルデヒドに変換される(図4参照)。

欠乏症
ヒトでの自然発生的なパントテン酸欠乏症は非常に稀で、重篤な栄養不良の場合にのみ観察されてきた。フィリピン、ミャンマー、および日本での第二次世界大戦捕虜は、足のしびれや苦痛を伴った灼熱感や刺痛を経験した。これらの症状は、特にパントテン酸補給で緩和された(4)。ヒトのパントテン酸欠乏症は、実験的にパントテン酸キナーゼ阻害薬(オメガ>メチルパントテン酸、上記の図1参照)とパントテン酸欠乏の食事を同時投与することで誘発されてきた。この実験の参加者は、頭痛、疲労、不眠、腸の不調、および手足のしびれやひりひり感を訴えた(12)。別の研究では、パントテン酸を含まない食事のみを与えられた参加者は欠乏症の臨床的兆候を示さなかったが、何人かはだるそうで疲労を訴えた(13)。
ホモパントテン酸カルシウム(ホパンテン酸カルシウム)は、コリン作動性効果(神経伝達物質のアセチルコリンの効果に似たもの)があるパントテン酸拮抗薬である。この化合物は日本で、特にアルツハイマー病の精神機能の強化に使用されている。これの稀な副作用は、肝臓が毒素を除去することができずに起こる異常な脳機能状態である肝性脳症の発症であった。肝性脳症はパントテン酸補給によって回復し、このことから肝性脳症はホモパントテン酸で誘発されたパントテン酸欠乏症であったことが示唆される(14)。留意したいのは、パントテン酸キナーゼII(上記の図1参照)を符号化するヒトの遺伝子であるPanK2の突然変異は、4'-ホスホパンテテインおよび補酵素Aの合成障害を起こすということである(「機能」の項参照)。パントテン酸キナーゼ関連神経変性症と呼ばれるこの障害は、視覚および知的障害、ジストニア(筋緊張異常)、言語異常、行動困難、および人格障害によって特徴付けられる(15)。
しかし、パントテン酸は自然に広く分布しておりヒトでの欠乏症は非常に稀であることから、欠乏症の結果に関するほとんどの情報は動物での実験研究から集められてきた((3)の文献でレビュー)。パントテン酸欠乏症のラットは副腎に損傷が起こり、サルはヘモグロビンの成分であるヘムの合成不足から貧血になった。パントテン酸欠乏症のイヌは低血糖、呼吸促迫や頻脈、および痙攣を起こした。ニワトリは皮膚刺激、羽毛の異常、およびミエリン鞘の変性に関連した脊髄神経損傷を発症した。パントテン酸が欠乏したマウスは運動耐性が下がり、筋肉や肝臓でのブドウ糖貯蔵(グリコーゲンの形態)が減るという症状を示した。マウスはまた、皮膚刺激や毛の白髪化も起こり、これはパントテン酸の投与で元に戻った。
症状が多様なことは、パントテン酸には補酵素の形態で多数の機能があることを強く示すものである。
目安量(AI)
ヒトでのパントテン酸の必要量に関する情報がほとんどないため、米国医学研究所の食品栄養委員会は健康な集団で観察された食事性摂取量に基づいて目安量(AI)を設定した(表1)(16)。
| 年齢層 | 年齢 | 男性(mg/日) | 女性(mg/日) |
|---|---|---|---|
| 乳児 | 0~6ヶ月 | 1.7 | 1.7 |
| 乳児 | 7~12ヶ月 | 1.8 | 1.8 |
| 子供 | 1~3歳 | 2 | 2 |
| 子供 | 4~8歳 | 3 | 3 |
| 子供 | 9~13歳 | 4 | 4 |
| 青少年 | 14~18歳 | 5 | 5 |
| 成人 | 19歳以上 | 5 | 5 |
| 妊娠期 | 全年齢 | - | 6 |
| 授乳期 | 全年齢 | - | 7 |
疾病治療
創傷治癒
D-パントテン酸カルシウムおよび/またはパントテノール(図5参照)を人工的に傷つけた皮膚線維芽細胞培養物の培地に添加すると細胞増殖や細胞移動が高まり、そのためin vitro(ガラス容器内)での創傷治癒が早くなることがわかった(17,18)。同様に、ガラス容器内でパントテン酸欠乏症状態を作ると、細胞増殖中の皮膚線維芽細胞の分化マーカーの発現を誘発し、ヒトのケラチノサイト(角化細胞)の増殖を阻害した(19)。D-パンテノールまたはデクスパンテノールとしても知られるD-パントテン酸カルシウムまたはパントテノールのどちらかを含む軟膏を皮膚に塗ると、動物の皮膚の創傷閉鎖を早め、瘢痕組織の強度が増すことが示されている(3)。
創傷治癒におけるデクスパンテノールの効果はよくわかっていない。12人の健康なボランティアを含むプラセボ対照研究で、デクスパンテノールを含む軟膏を皮膚の創傷治癒モデルに塗った(12時間ごとに1~6日間)ら、細胞増殖、炎症、および組織修復のマーカーの発現が強化されるという関連があった(20)。しかしこの研究では、デクスパンテノールの局所適用に反応したこれらの変化がプラセボに比べて傷の修復プロセスを向上させたのかどうかを報告していない(20)。何の効果も示されなかった研究もある。タトゥー消去の手術を受けた患者による初期の無作為化対照試験で、1または3グラムのビタミンCと200mgまたは900mgのパントテン酸を21日間毎日併用補給しても、創傷治癒プロセスはあまり向上しなかったことがわかった(21,22)。しかし最近の無作為化二重盲検プラセボ対照研究で、デクスパンテノールのトローチ(300mg/日を最長で術後14日間)の使用によって、扁桃摘出術を受けた子供の粘膜治癒が早くなったことがわかった(23)。
高コレステロール
初期の研究で、パンテノン酸の誘導体であるパンテチンを薬理学的用量で使用すると、コレステロール低減効果があるかもしれないことが示唆された(24,25)。パンテチンは2つのパンテチン分子がジスルフィド結合(2つの硫黄分子間の化学結合)によって結合してできている(図5参照)。パンテチンは補酵素Aに構造的に関連があり、アシルキャリアタンパク質であるホルミルテトラヒドロ葉酸脱水素酵素の生物学的機能に必要な補欠分子族と、α-アミノアジピン酸セミアルデヒド合成酵素に見られる(「機能」の項参照)。16週間の無作為化二重盲検プラセボ対照研究で、毎日のパンテチン補給(600mg/日を8週間、その後900mg/日を8週間)によって、心血管疾患(CVD)リスクが低~中の120人の脂質パラメータプロフィールが大きく向上した。ベースライン(試験開始時)への調整の後で、低密度リポタンパク質コレステロール(LDL-C)およびアポリポタンパク質B(apoB)濃度を低下させること、およびトリグリセリド(中性脂肪)と高密度リポタンパク質コレステロールの比率(TG:HDL-C)を下げることにおいて、パンテチンはプラセボよりもかなり効果的であったことがわかった(26)。パンテチンは耐容性がよくコレステロール代謝を向上させるのにも有益な可能性があるようだが、パンテチンはビタミンではない。薬理学的用量のパンテチンを高い血中コレステロールやトリグリセリドの治療に使用するという決定は、適切な追跡をすることができる有資格医療提供者が合意した場合のみなされねばならない。

白髪化
パントテン酸欠乏のマウスは皮膚刺激や毛が白くなる症状を起こしたが、これはパントテン酸の投与で元に戻った。ヒトでは、パントテン酸をサプリメントとして摂取したりパントテン酸入りのシャンプーを使用したりすることが髪の色を保ったり復元したりすることができるというエビデンスはない。
摂取源
食品の摂取源
パントテン酸は、通常は補酵素A(CoA)や4'-ホスホパンテテインの成分として様々な食物に含まれる(上記の図1参照)。消化の際には、食事性の補酵素Aとホスホパンテテインは腸での吸収の前にパントテン酸に加水分解される(3)。動物の肝臓や腎臓、魚、貝、豚肉、鶏肉、卵黄、牛乳、ヨーグルト、豆類、マッシュルーム、アボカド、ブロッコリー、およびサツマイモは、パントテン酸の良好な摂取源である。穀物もパントテン酸の良好な摂取源であるが、加工や精製することで35~70%が喪失しかねない。冷凍や缶詰も同様の喪失が起こる(16)。大規模な国家的栄養調査では、パントテン酸摂取量の推定ができなかった。これは主に食物のパントテン酸含有量データが不足していたからである(16)。小規模研究では、パントテン酸の毎日の平均摂取量は成人で4~7mgと推定された。表2はパントテン酸の豊富な食物とその含有量をミリグラムで表示したものである。食物の栄養素含有量の詳細情報は、USDAの食品成分データベースを検索のこと。
腸内細菌
結腸(大腸)に通常棲み着いている細菌は、パントテン酸を合成可能である。ビオチンとパントテン酸の摂取のための特別な輸送体が結腸内壁の培養細胞において同定されたことから、ヒトは腸内細菌によって生成されたパントテン酸とビオチンを吸収できるのではないかと示唆された(27)。しかし、細菌による合成がヒトのパントテン酸摂取にどの程度貢献しているかは不明である(28)。
サプリメント
パントテノールおよびパントテン酸塩
サプリメントは一般的にパントテン酸の安定したアルコール類似体であるパントテノール(パンテノール)を含み、これはヒトでは急速にパントテン酸に変換される。パントテン酸のカルシウムおよびナトリウム塩であるD-パントテン酸カルシウムおよびナトリウムも、サプリメントとして市販されている。
パンテチン
パンテチンは日本でコレステロール低減薬として使用されており、米国では栄養補助食品として市販されている(29)。
安全性
毒性
パントテン酸はヒトに有毒であるとはされていない。注目された唯一の有害作用は、D-パントテン酸カルシウムを10~20g/日という非常に高用量で摂取した結果としての下痢である(30)。しかし、10mg/日のビオチンと300mg/日のパントテン酸を2ヶ月間併用した年配女性が、命に関わる好酸球性胸膜心膜滲出(好酸球性胸水)を起こした報告が1つある(31)。パントテン酸の食事摂取基準量(DRI)が1998年に確定された際には有害作用の報告がなかったため、米国医学研究所の食品栄養委員会はパントテン酸の許容上限摂取量(UL)を設定しなかった(16)。パンテチンは最大1,200mg/日まで一般的によく耐用性がある。しかし、吐き気や胸焼けなどの胃腸系の副作用が報告されている(29)。また、デクスパンテノール(D-パンテノール)を最大5%まで含む局所製剤は、最長1ヶ月まで安全に使用されてきた。それでもデクスパンテノール含有の軟膏の使用による皮膚刺激、接触性皮膚炎、および湿疹の報告がいくつかある(32,33)。
栄養素相互作用
大用量のパントテン酸は、ヒトのナトリウム依存性マルチビタミン輸送体(hSMVT)による腸および細胞での取り込みにおいて、ビオチンと競合する可能性がある(27,34)。
薬物相互作用
エストロゲンやプロゲスチンを含む経口避妊薬は、パントテン酸の需要を増やすかもしれない(30)。スタチン(HMG-CoA還元酵素阻害薬)と呼ばれるコレステロール低減薬またはニコチン酸 とパンテチンを併用すると、血中脂質に相加効果をもたらすかもしれない(29)。
ライナス・ポーリング研究所の推奨
健康を最大限促進し慢性疾患を防ぐために必要な食事性パントテン酸の量を決めるためのさらなるデータが必要である。ライナス・ポーリング研究所は、食品栄養委員会が設定した成人に対する5mg/日というパントテン酸の推奨量を支持する。様々な食事によって、十分なパントテン酸がほとんどの人には提供されているはずである。パントテン酸の一日摂取量(DV)を100%含むマルチビタミン/ミネラルのサプリメントを毎日摂取するというライナス・ポーリング研究所の推奨に従えば、少なくとも5mg/日の摂取が確実であろう。
年配者(50歳超)
パントテン酸の摂取または必要量が年配者では異なるというエビデンスは、現在ほとんどない。たいていのマルチビタミン/ミネラルのサプリメントで、少なくとも5mg/日のパントテン酸が摂れる。
Authors and Reviewers
Written in May 2004 by:
Jane Higdon, Ph.D.
Linus Pauling Institute
Oregon State University
Updated in April 2008 by:
Victoria J. Drake, Ph.D.
Linus Pauling Institute
Oregon State University
Updated in April 2015 by:
Barbara Delage, Ph.D.
Linus Pauling Institute
Oregon State University
Reviewed in July 2015 by:
Robert B. Rucker, Ph.D.
Distinguished Professor Emeritus
Department of Nutrition and School of Medicine
University of California, Davis
Copyright 2000-2026 Linus Pauling Institute
Figure Alternative Text
Figure 1. Coenzyme A synthesis from pantothenic acid
The figure illustrates the biochemical pathway that converts pantothenic acid (pantothenate) into coenzyme A, showing chemical structures of intermediates and the enzymes involved.
At the top of the figure, pantothenic acid is phosphorylated by pantothenic acid kinase 2 (PANK2) using ATP, and releasing ADP, to form 4’-phosphopantothenic acid. This step is impaired in individuals with PANK2 gene mutations.
Next, 4’-phosphopantothenic acid is converted to 4’-phosphopantothenoylcysteine with 4-phosphopantothenic acid synthetase requiring CTP and cysteine and releasing CDP and inorganic phosphate.
Then 4’-phosphopantothenic acid decarboxylase removes CO2 to produce 4’-phosphopantetheine.
Using ATP, dephospho-CoA pyrophosphorylase converts 4’-phosphopantetheine into 4’-dephospho-coenzyme A, releasing pyrophosphate.
Finally, dephospho-CoA kinase phosphorylates 4’-dephospho-coenzyme using ATP, forming coenzyme A and releasing ADP.
The figure also shows the CoA hydrolase reaction, which converts coenzyme A back into 4’-phosphopantetheine, releasing 3’,5’-ADP.
Figure 2. 4’-Pantetheinylation of acyl-carrier protein (ACP)
The figure illustrates the post-translational modification of ACP called 4’-pantetheinylation. This process adds a 4’-phosphopantetheinyl moiety, a prosthetic group required for ACP’s biological activity.
The enzyme 4’-phosphopantetheinyl transferase catalyzes the hydrolysis of coenzyme A to 3’,5’-adenosine diphosphate and 4’-phosphopantetheine, and the transfer of the 4’-phosphopantetheinyl moiety to a serine residue at the active site of ACP, converting biologically inactive apo-ACP into active holo-ACP.
Figure 3. 4’-Pantetheinylation of formyltetrahydrofolate dehydrogenase (FDH)
The figure shows two reactions involving FDH:
Panel A: Holo-FDH (the biologically active FDH) catalyzes the conversion of 10-formyltetrahydrofolate into tetrahydrofolate using water and NADP+, releasing CO2, NADPH, and a proton (H+). FDH requires a prosthetic group, a 4’-phosphopantetheinyl moiety, for biological activity.
Panel B: In a post-translational modification called 4’-pantetheinylation, the enzyme 4’-phosphopantetheinyl transferase catalyzes the hydrolysis of coenzyme A into 3’,5’-adenosine diphosphate and 4’-phosphopantetheine and the transfer of the 4’-phosphopantetheinyl moiety to a specific serine residue of FDH. This converts biologically inactive apo-FDH to active holo-FDH.
Figure 4. 4’-Pantetheinylation of α-aminoadipate semialdehyde synthase (AASS)
The figure illustrates the mitochondrial enzyme AASS that has two domains: the lysine-ketoglutarate reductase domain and the saccharopine dehydrogenase domain. These domains catalyze two reactions once AASS is biologically active (holo-AASS).
The main part of the figure shows 4’-pantetheinylation of AASS, the post-translational modification required for its biological activity. 4’-Phosphopantetheinyl transferase catalyzes the transfer of a 4’-phosphopantetheinyl moiety from coenzyme A to a specific residue of AASS, converting apo-AASS to holo-AASS.
First reaction: Holo-AASS converts lysine to saccharopine, using NADPH, a proton (H+), and α-ketoglutarate, releasing H2O and NADP+.
Second reaction: Holo-AASS converts saccharopine into α-aminoadipate semialdehyde using H2O and NAD+, releasing NADH, H+, and glutamate.
Figure 5. Chemical structures of some pantothenic acid derivatives
The figure shows the chemical structures of the pantothenic acid derivatives: pantothenol (panthenol), calcium pantothenate, and pantethine.
References
- Trumbo PR. Pantothenic acid. In: Ross AC, Caballero B, Cousins RJ, Tucker KL, Ziegler TR, eds. Modern Nutrition in Health and Disease. 11th ed. Baltimore: Lippincott Williams & Wilkins; 2014:351-357.
- Martinez DL, Tsuchiya Y, Gout I. Coenzyme A biosynthetic machinery in mammalian cells. Biochem Soc Trans. 2014;42(4):1112-1117. (PubMed)
- Miller JW, Rucker RB. Pantothenic acid. In: Erdman JWJ, Macdonald IA, Zeisel SH, eds. Present Knowledge in Nutrition. 10th ed. Ames: Wiley-Blackwell; 2012:375-390.
- Bauerly K, Rucker RB. Pantothenic acid. In: Zempleni J, Rucker RB, McCormick DB, Suttie JW, eds. Handbook of vitamins. 4th ed. Boca Raton: CRC Press; 2007:289-314.
- Takahashi A, Mizusawa K. Posttranslational modifications of proopiomelanocortin in vertebrates and their biological significance. Front Endocrinol (Lausanne). 2013;4:143. (PubMed)
- Choudhary C, Weinert BT, Nishida Y, Verdin E, Mann M. The growing landscape of lysine acetylation links metabolism and cell signalling. Nat Rev Mol Cell Biol. 2014;15(8):536-550. (PubMed)
- Beld J, Sonnenschein EC, Vickery CR, Noel JP, Burkart MD. The phosphopantetheinyl transferases: catalysis of a post-translational modification crucial for life. Nat Prod Rep. 2014;31(1):61-108. (PubMed)
- Bunkoczi G, Pasta S, Joshi A, et al. Mechanism and substrate recognition of human holo ACP synthase. Chem Biol. 2007;14(11):1243-1253. (PubMed)
- Donato H, Krupenko NI, Tsybovsky Y, Krupenko SA. 10-formyltetrahydrofolate dehydrogenase requires a 4'-phosphopantetheine prosthetic group for catalysis. J Biol Chem. 2007;282(47):34159-34166. (PubMed)
- Strickland KC, Hoeferlin LA, Oleinik NV, Krupenko NI, Krupenko SA. Acyl carrier protein-specific 4'-phosphopantetheinyl transferase activates 10-formyltetrahydrofolate dehydrogenase. J Biol Chem. 2010;285(3):1627-1633. (PubMed)
- Strickland KC, Krupenko NI, Dubard ME, Hu CJ, Tsybovsky Y, Krupenko SA. Enzymatic properties of ALDH1L2, a mitochondrial 10-formyltetrahydrofolate dehydrogenase. Chem Biol Interact. 2011;191(1-3):129-136. (PubMed)
- Hodges RE, Ohlson MA, Bean WB. Pantothenic acid deficiency in man. J Clin Invest. 1958;37(11):1642-1657. (PubMed)
- Fry PC, Fox HM, Tao HG. Metabolic response to a pantothenic acid deficient diet in humans. J Nutr Sci Vitaminol (Tokyo). 1976;22(4):339-346. (PubMed)
- Bender DA. Optimum nutrition: thiamin, biotin and pantothenate. Proc Nutr Soc. 1999;58(2):427-433. (PubMed)
- Kurian MA, Hayflick SJ. Pantothenate kinase-associated neurodegeneration (PKAN) and PLA2G6-associated neurodegeneration (PLAN): review of two major neurodegeneration with brain iron accumulation (NBIA) phenotypes. Int Rev Neurobiol. 2013;110:49-71. (PubMed)
- Food and Nutrition Board, Institute of Medicine. Pantothenic acid. Dietary Reference Intakes: Thiamin, Riboflavin, Niacin, Vitamin B6, Vitamin B12, Pantothenic Acid, Biotin, and Choline. Washington, D.C.: National Academy Press; 1998:357-373. (National Academy Press)
- Weimann BI, Hermann D. Studies on wound healing: effects of calcium D-pantothenate on the migration, proliferation and protein synthesis of human dermal fibroblasts in culture. Int J Vitam Nutr Res. 1999;69(2):113-119. (PubMed)
- Wiederholt T, Heise R, Skazik C, et al. Calcium pantothenate modulates gene expression in proliferating human dermal fibroblasts. Exp Dermatol. 2009;18(11):969-978. (PubMed)
- Kobayashi D, Kusama M, Onda M, Nakahata N. The effect of pantothenic acid deficiency on keratinocyte proliferation and the synthesis of keratinocyte growth factor and collagen in fibroblasts. J Pharmacol Sci. 2011;115(2):230-234. (PubMed)
- Heise R, Skazik C, Marquardt Y, et al. Dexpanthenol modulates gene expression in skin wound healing in vivo. Skin Pharmacol Physiol. 2012;25(5):241-248. (PubMed)
- Vaxman F, Olender S, Lambert A, et al. Effect of pantothenic acid and ascorbic acid supplementation on human skin wound healing process. A double-blind, prospective and randomized trial. Eur Surg Res. 1995;27(3):158-166. (PubMed)
- Vaxman F, Olender S, Lambert A, Nisand G, Grenier JF. Can the wound healing process be improved by vitamin supplementation? Experimental study on humans. Eur Surg Res. 1996;28(4):306-314. (PubMed)
- Celebi S, Tepe C, Yelken K, Celik O. Efficacy of dexpanthenol for pediatric post-tonsillectomy pain and wound healing. Ann Otol Rhinol Laryngol. 2013;122(7):464-467. (PubMed)
- Coronel F, Tornero F, Torrente J, et al. Treatment of hyperlipemia in diabetic patients on dialysis with a physiological substance. Am J Nephrol. 1991;11(1):32-36. (PubMed)
- Gaddi A, Descovich GC, Noseda G, et al. Controlled evaluation of pantethine, a natural hypolipidemic compound, in patients with different forms of hyperlipoproteinemia. Atherosclerosis. 1984;50(1):73-83. (PubMed)
- Rumberger JA, Napolitano J, Azumano I, Kamiya T, Evans M. Pantethine, a derivative of vitamin B(5) used as a nutritional supplement, favorably alters low-density lipoprotein cholesterol metabolism in low- to moderate-cardiovascular risk North American subjects: a triple-blinded placebo and diet-controlled investigation. Nutr Res. 2011;31(8):608-615. (PubMed)
- Said HM, Ortiz A, McCloud E, Dyer D, Moyer MP, Rubin S. Biotin uptake by human colonic epithelial NCM460 cells: a carrier-mediated process shared with pantothenic acid. Am J Physiol. 1998;275(5 Pt 1):C1365-1371. (PubMed)
- Said HM. Intestinal absorption of water-soluble vitamins in health and disease. Biochem J. 2011;437(3):357-372. (PubMed)
- Hendler SS, Rorvik DR, eds. PDR for Nutritional Supplements. 2nd ed. Montvale: Thomson Reuters; 2008.
- Flodin N. Pharmacology of micronutrients. New York: Alan R. Liss, Inc.; 1988.
- Debourdeau PM, Djezzar S, Estival JL, Zammit CM, Richard RC, Castot AC. Life-threatening eosinophilic pleuropericardial effusion related to vitamins B5 and H. Ann Pharmacother. 2001;35(4):424-426. (PubMed)
- Herbst RA, Uter W, Pirker C, Geier J, Frosch PJ. Allergic and non-allergic periorbital dermatitis: patch test results of the Information Network of the Departments of Dermatology during a 5-year period. Contact Dermatitis. 2004;51(1):13-19. (PubMed)
- Schmuth M, Wimmer MA, Hofer S, et al. Topical corticosteroid therapy for acute radiation dermatitis: a prospective, randomized, double-blind study. Br J Dermatol. 2002;146(6):983-991. (PubMed)
- Chirapu SR, Rotter CJ, Miller EL, Varma MV, Dow RL, Finn MG. High specificity in response of the sodium-dependent multivitamin transporter to derivatives of pantothenic acid. Curr Top Med Chem. 2013;13(7):837-842. (PubMed)
免責事項
ライナスポーリング研究所微量栄養素情報センターは、一般向けの食事要因、健康面、栄養補助食品、食品および飲料に関する科学的情報を提供する。著者および出版社は、本サイト上の医学的、心理学的、栄養学的カウンセリングサービスを利用しないことを合意した上で、本情報を利用することができる。本情報は、有資格の医療従事者または栄養士への相談の代わりとして利用することはできない。
本ウェブサイトに掲載されている食事要因、栄養補助食品、食品および飲料に含まれる情報は、実施可能なすべての使用法、行為、使用上の注意、副作用、および相互作用を網羅するものではない。本ウェブサイトは、個々の問題についての栄養学的、医学的助言をすることは意図していない。本サイトの電子的に利用可能な情報に基づく個人的な行為、または不作為に対する責任は、明示的に否認される。
本ウェブサイトにおける著作権を有する資料のいずれも、複製、修正、配布、表示、発信、実行、公開または販売をおこなってはならない。本ウェブサイトにハイパーリンクを貼ることは可能であるが、その際には以下の文言を付記することとする。
「本リンクは、オレゴン州立大学ライナスポーリング研究所の提供するウェブサイトへ移動します。<<名前>>は、オレゴン州立大学ライナスポーリング研究所のアフィリエイトではなく、認証されたリンクでもありません」